




| Research Article | ||
Open Vet. J.. 2026; 16(5): 2601-2609 Open Veterinary Journal, (2026), Vol. 16(5): 2601-2609 Research Article Different dose-based multi HN&F epitope mRNA vaccine against Newcastle diseaseAsma Abdlhusen Shrshoh* and Nafea Sabih JasimDepartment of Pathology and Poultry Diseases, College of Veterinary Medicine, University of Al-Qadisiyah, Al-Qādisiyyah, Iraq *Corresponding Author: Asma Abdlhusen Shrshoh. Department of Pathology and Poultry Diseases, College of Veterinary Medicine, University of Al-Qadisiyah, Al-Qādisiyyah, Iraq. Email: vet.post24.6 [at] qu.edu.iq Submitted: 28/01/2026 Revised: 17/03/2026 Accepted: 25/03/2026 Published: 31/05/2026 © 2025 Open Veterinary Journal
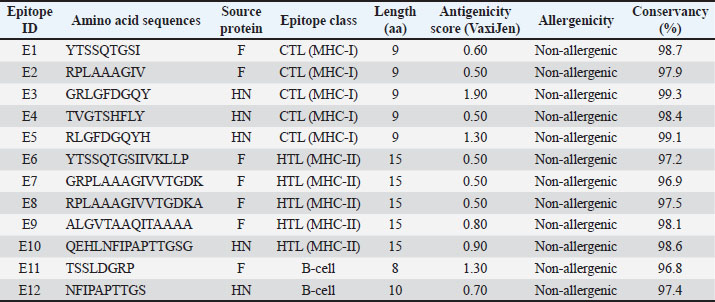
AbstractBackground: Newcastle disease virus (NDV) is considered a major player in the ongoing challenges in the poultry farming system due to its virulence, which affects productivity. Aim: This study aimed to develop and evaluate a multi-epitope mRNA vaccine that targets the NDV fusion (F) and hemagglutinin–neuraminidase (HN) proteins in chickens. Methods: The cytotoxic T lymphocyte (CTL) epitopes were obtained from NetCTL 1.2, whereas the helper T lymphocyte (HTL) epitopes were predicted using the IEDB MHC-II binding prediction tool. The subsequent selection of the epitopes was via binding affinity and was also predicted antigenically (VaxiJen v2.0) and allergenically (AllerTOP v2.0), and for sequence conservancy. The selected epitopes were fused with Alanine–Alanine–Tyrosine, Glycine–Proline–Glycine–Proline–Glycine, and Lysine–Lysine linkers. The vector was constructed, and the sequence was validated using VectorBee. There were 280 chickens distributed randomly across eight sets (n=35 per set), consisting of one set that received the F&HN multi-epitope mRNA vaccine in addition to one set that was unvaccinated and served as a negative control, and six sets that received one of several different commercially available NDV vaccines. A virulent NDV strain was used to challenge the birds, and hemagglutination inhibition (HI), ELISA, lymphocyte proliferation, and Interferon-gamma Enzyme-Linked ImmunoSpot assays were used to assess the immune responses. Results: Several epitopes for CTL, HTL, and B-cells that were both conservatively positioned and highly immunogenic were identified for the F and HN proteins. In the experiment, chickens that were immunized with the multi-epitope mRNA vaccine had significantly higher HI antibody titers and ELISA optical density values than the negative control group. The mRNA vaccine group had statistically significant differences in lymphocyte proliferation and the number of IFN-γ-producing cells. After the virulent NDV challenge, the mRNA vaccine group had the best protection with the fewest and least severe clinical signs and the lowest mortality. Conclusion: The mRNA vaccine for the F and HN multi-epitopes NDV was the first to provide extensive protection against NDV. The vaccine elicited strong immune responses, demonstrating the potential of mRNA multi-epitope-based vaccines for NDV as a significant improvement over the previous method. Keywords: Antigenicity, Fusion protein, mRNA vaccine, Newcastle disease virus, T-cell epitopes. IntroductionNewcastle disease virus (NDV) is very contagious and continues to threaten the poultry industry worldwide, mainly in areas affected by the virulent genotype VII strains. Owing to antigenic divergence, poor vaccine coverage, and low cell-mediated immunity (Sun et al., 2024; Wu et al., 2025), outbreaks occur even when inactivated and live-attenuated vaccines are widely used. Most NDV vaccines focus on the elicitation of a humoral immune response, whereas the activation of cytotoxic T lymphocytes (CTL) and helper T cell responses is variable. This is likely responsible for the incomplete protection and subsequent viral shedding after the challenge (Pandarangga et al., 2024). Modern technology in immunoinformatics has enabled the creation of multi-epitope vaccines that activate various immune responses and target conserved areas of viruses. Several in silico studies have shown that vaccines targeting the structural proteins of NDV, especially fusion (F) and hemagglutinin–neuraminidase (HN), have great coverage in terms of population and antigenicity (Tataje-Lavanda et al., 2023; Zeb et al., 2024; Randriamamisolonirina et al., 2025). They are ideal candidates for neutralizing antibody responses and cell-mediated immune responses, given their importance in viral cell F and membrane regulation (Mozafari et al., 2022; Ji et al., 2025). However, most NDV vaccines based on epitope technology are limited to vector-based and peptide-based platforms, and few have looked into current nucleic acid-based vaccine platforms. mRNA vaccines are safe, rapidly developed, and effective at stimulating both humoral and cellular immunity. Vaccines using modified untranslated regions, codon use, and epitope structure have been shown to improve the expression and immunogenicity of individual antigens in the avian cellular system (Chowdhury et al., 2024; Pandarangga et al., 2024). Furthermore, the use of a single mRNA construct that expresses multiple epitopes may solve the problems of strain variability and suboptimal immune response of conventional NDV vaccines (Kamran et al., 2026). In this regard, this study focused on designing, constructing, and assessing the immunogenicity and protective efficacy of a multi-epitope mRNA vaccine. This vaccine was developed to encode the conserved B-cell, CTL, and helper T lymphocyte (HTL) epitopes from NDV F and HN proteins and to evaluate its efficacy against commercially available NDV vaccines in chickens. Materials and MethodsObtaining and analyzing the sequence conservationUsing the National Center for Biotechnology Information database, the full-length amino acid sequences of the F and HN proteins of NDV Genotype VII, in Fast-All sequence comparison format format, were retrieved. Considering confirmed classification and relevance to circulating virulent strains, representative Genotype VII sequences were selected. The default alignment parameters were used to conduct multiple sequence alignments in Clustal Omega (EMBL-EBI, United Kingdom) to determine the conserved sequences within the retrieved sequences. To ensure maximal protective coverage, segments with a high degree of conservancy and low sequence variability were chosen to proceed to the next step of epitope prediction. Prediction of immunogenic epitopesThe linear B-cell epitopes were predicted on the BepiPred 2.0 server (Technical University of Denmark) using a default threshold of 0.5, which is used to detect linear regions of antigens. For the conformational B-cell epitopes, Immune Epitope Database (IEDB) Analysis Resource ElliPro tools are used based on 3D-conformation of the protein and protrusion index scoring. The NetCTL 1.2 server (DTU Bioinformatics, Denmark) was used to predict the CTL epitopes, which combines proteasome cleavage prediction, antigen processing and transport efficiency, and Major Histocompatibility Complex class I (a) molecule binding prediction (a score of 0.75 was used as a threshold). The MHC-II binding prediction tool from IEDB was used to predict the HTL epitopes based on the binding prediction consensus approach. Epitopes with low percentiles, hence high predicted binding affinity, and to MHC-II (a) from chicken were chosen for further analyses. Antigenicity, allergenicity, and conservancy evaluationUsing the VaxiJen v2.0 server with the virus model and a threshold value of 0.4, all predicted epitopes were examined for antigenicity. Epitopes predicted to be antigenic were evaluated for allergenicity using AllerTOP v2.0, and only those epitopes that were classified as non-allergenic were retained. The IEDB Epitope Conservancy Tool was used to verify the extent of epitope conservancy among the NDV genotype VII strains, thereby ensuring the coverage of various strains and minimizing the potential for immune escape.2.4 Design and Codon Optimization of Multi-Epitope mRNA Constructs Immunologically optimized linkers were used to construct a single open reading frame for CTL, HTL, and B-cell epitopes. These included Alanine–Alanine–Tyrosine (AAY) for CTL epitopes to aid in proteasome processing, Glycine–Proline–Glycine–Proline–Glycine (GPGPG) for HTL to facilitate MHC-II binding, and Lysine–Lysine (KK) for B-cell epitopes to retain conformational flexibility. The mRNA construct design included a 5' Cap 1, a Kozak sequence (GCCACC), and a 3' poly(A) tail of 120 adenine nucleotides to improve mRNA stability and translation efficiency. The GenSmart Codon Optimization Tool (GenScript Biotech, USA) was used to perform codon optimization for Gallus gallus (chicken) expression. The codon adaptation index (CAI), guanine-cytosine (GC) content, and frequency of rare codons were considered. The integrity of the construct, reading frame, and sequence orientation was checked using the VectorBee software (CyberGene AB, Sweden). In vitro transcription and isolation of mRNAAn optimized coding sequence was inserted into the T7 promoter vector for in vitro transcription with the HiScribe™ T7 ARCA mRNA Kit with tailing (Catalog No. E2065S, New England Biolabs, USA). Each transcription reaction contained one inclusive microgram of DNA template in a final volume of 20 l and was incubated for 2 hours at 37°C according to the manufacturer’s instructions. The synthesized mRNA was capped and polyadenylated and then purified with the MEGAclear™ Transcription Clean-Up Kit (Catalog No. AM1908, Thermo Fisher Scientific, USA). mRNA was concentrated, and its purity was measured using a NanoDrop™ 2000 spectrophotometer (Thermo Fisher Scientific, USA). Source and housing of the test animalsSource and housing of 280 specific-pathogen-free broiler chickens were obtained from a certified commercial hatchery. They were placed in animal facilities with biosafety level 2. The environmental conditions were controlled at a temperature of 22℃–25℃ with a relative humidity of 50%–60%, and a 12-hour light and dark cycle. During the trial, the birds had ad libitum access to standard commercial feed and water. Vaccination protocol and experimental groupsAn experimental design framework was developed to measure the added value of the multi-epitope mRNA vaccine against the standard vaccination methods for Newcastle disease. To minimize selection bias and enhance biological uniformity, 280 specific-pathogen-free broiler chickens were equally divided into eight experimental groups (n=35 birds per group). One group was given the experimental mRNA vaccine, one group was assigned an unvaccinated negative control, and six groups were provided with commercially available NDV vaccines (field route vaccines, eye drops, drinking water, or inactivated formulations). This design replicated the vaccination routines practiced in the poultry farms of the study area and provided the opportunity for the experimental population to emulate the vaccination programs for Newcastle disease in field poultry populations. The chickens were randomly allocated to eight experimental groups. Each experimental group consisted of 35 birds. The first group was vaccinated with the multi-epitope mRNA vaccine, which was administered intramuscularly into the breast muscle at 20 µg per bird. The vaccine was diluted in 100 µl of sterile phosphate-buffered saline (PBS, pH 7.4) and injected using sterile 1 ml syringes. The second group was an unvaccinated negative control and received PBS only. The other six groups received Newcastle Disease Vaccines, which were administered via spray, eye-drop, drinking water, or inactivated formulations, in accordance with the manufacturers’ recommendations. Virulent NDV challengeAll the birds were challenged with the virulent NDV Genotype VII strain 28 days post-vaccination at a dose of 10⁶ EID₅₀ per bird via the oculo-nasal route. Birds were monitored for 14 consecutive days post-challenge to capture clinical signs and record morbidity and mortality. Assessment of humoral immune responsesBlood samples were collected, and serum was separated for each time point to evaluate the humoral immune response. Hemagglutination inhibition (HI) assays were performed using standard NDV antigen containing four hemagglutinating units as per OIE specifications following the standard heat inactivation procedure for serum samples, which involves heating to 56℃ for 30 minutes. HI titers were recorded in log₂ format. NDV-specific antibodies were quantified using a commercial enzyme-linked immunosorbent assay kit (Catalog No. CK119, IDEXX Laboratories, USA), and optical density (OD) was read at 450 nm using a microplate reader as per the manufacturer’s instructions. Cell-mediated immune response evaluationPeripheral Blood Mononuclear Cells were isolated from whole blood using Ficoll-Paque™ PLUS (Catalog No. 17-1440-03, Cytiva, Sweden) density gradient medium. The 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay (Catalog No. M5655, Sigma-Aldrich, USA), a colorimetric assay for assessing cell proliferation, was used, and the results were expressed in terms of the stimulation index (SI). Cells producing interferon-gamma were measured using a Chicken Interferon-gamma Enzyme-Linked ImmunoSpot (IFN-γ ELISPOT) kit (Catalog No. 3130-2A, Mabtech, Sweden), and the spots were counted using an automatic ELISPOT reader. Statistical analysisStatistical analysis of experimental data was performed using GraphPad Prism version 9.5.1 (GraphPad Software, San Diego, CA). Data are expressed as mean standard error. Comparisons among groups were performed using one-way analysis of variance, followed by Tukey’s post hoc test. A p-value of 0.05 was considered statistically significant. Ethical approvalThe Committee for Research Ethics at the College of Veterinary Medicine, University of Al-Qadisiyah, Iraq, approved this study. ResultsSelection and quantitative analysis of F and HN epitopes using immunoinformatics analysisImmunoinformatics analysis led to the selection of 22 epitopes representing the F and HN proteins of NDV genotype VII. These 22 epitopes included 13 for CTL, 5 for HTL, and 4 for B cells. Using VaxiJen v2.0, which calculates provisional antigenicity scores, F-derived epitopes ranged from 0.50 to 1.90, yielding an average of 0.71 ± 0.06, and HN-derived epitopes averaged 0.74 ± 0.05. Conservancy analysis of the selected F and HN proteins across the queried genotype VII strains revealed that 19 of the 22 epitopes (86.4%) had a sequence conservancy of 98% and higher. The conservancy for the remaining epitopes ranged from 96.8% to 97.5%. All selected epitopes were classified as non-allergenic by AllerTOP v2.0 (Table 1). Table 1. Immunoinformatics-based quantitative characteristics of selected F and HN epitopes included in the vaccine.
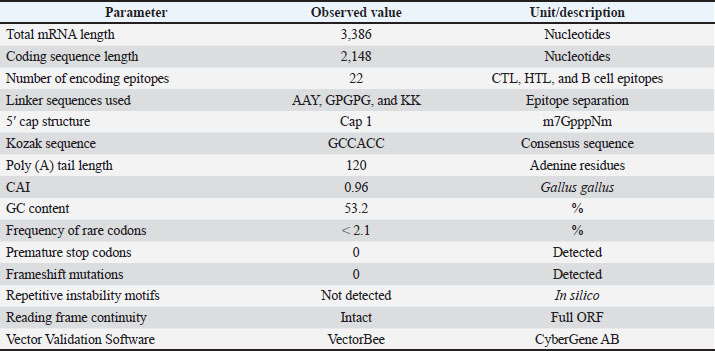
Molecular characterization and integrity of the multi-epitope mRNA constructThe complete multi-epitope mRNA construct, including the optimized coding sequence and its accompanying regulatory elements, was 3,386 nucleotides long. After optimizing the coding sequence for the target species, G. gallus, the corresponding CAI was 0.96, and the GC content was 53.2%, both of which are considered optimal for expression in the avian cellular system. The sequence was found to be clear of the following undesirable features: premature termination codons, frame shifts, and other sequence instability indications. The in-silico assessments also verified that the mRNA construct contained no interruptions in the open reading frames and maintained a poly(A) tail of 120 nucleotides (Table 2). Table 2. Molecular composition and integrity parameters of the designed multi-epitope mRNA construct.
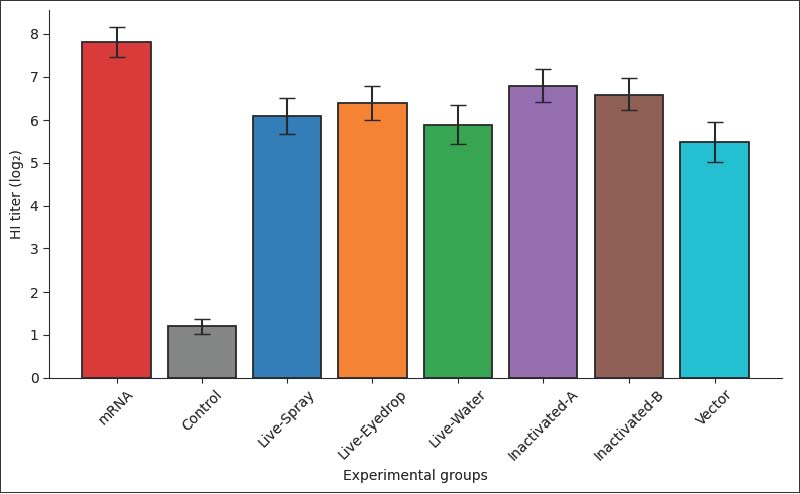
Humoral immune responses measured by HI and ELISAHI antibody titers at 28 days post-vaccination indicate variability within the experimental groups. The multi-epitope mRNA vaccine group had the highest mean HI titer at 7.8 ± 0.35 log₂, the control unvaccinated group had a mean HI titer of only 1.2 ± 0.18 log₂ (p < 0.001). The commercially vaccinated groups also had significantly higher HI titers than the control groups (p < 0.01), and HI titers in the commercially vaccinated groups ranged from 5.5 ± 0.47 to 6.8 ± 0.38 log₂. ELISA results for mRNA vaccine groups, controls, and commercial vaccine groups had mean OD of 1.42 ± 0.09, 0.21 ± 0.04, and 0.96 ± 0.08 to 1.18 ± 0.07, respectively. The mRNA vaccine group also exhibited a coefficient of variation of <10%, indicating less variability compared with the other groups (Fig. 1).
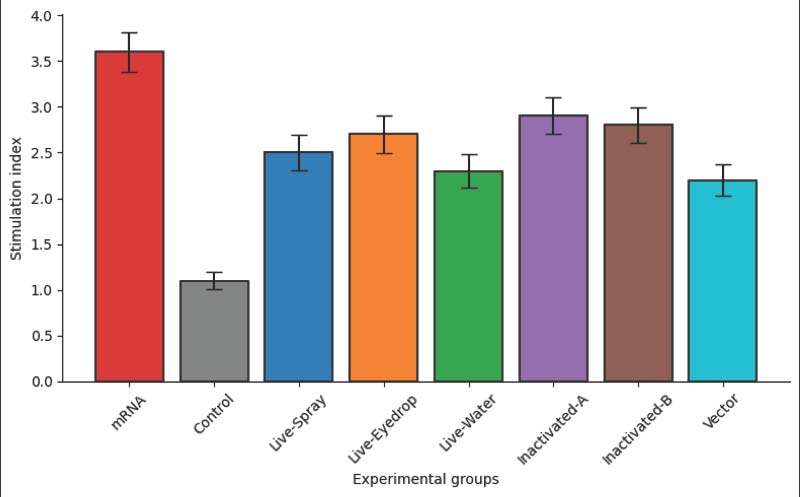
Fig. 1. HI antibody titers measured 28 days after vaccination in eight experimental groups. Bars represent mean log₂ HI titers, and error bars indicate the standard error. Cell-mediated immune responsesLymphocyte proliferation assays indicated that the mRNA vaccine group had a SI of 3.6 ± 0.22 compared to 1.1 ± 0.09 in the control group (p < 0.001). The SI of the commercial vaccine groups were between 2.3 ± 0.19 and 2.9 ± 0.21, and all were significantly different from those of the control group (p < 0.01). In the IFN-γ ELISPOT assay, the mRNA vaccine group had 286 ± 24 spot-forming units (SFU)/10⁶ cells, whereas the control group had 34 ± 8 SFU/10⁶ cells (p < 0.001). The commercial vaccine groups had 142 ± 17 to 214 ± 20 SFU/10⁶ cells (Fig. 2).
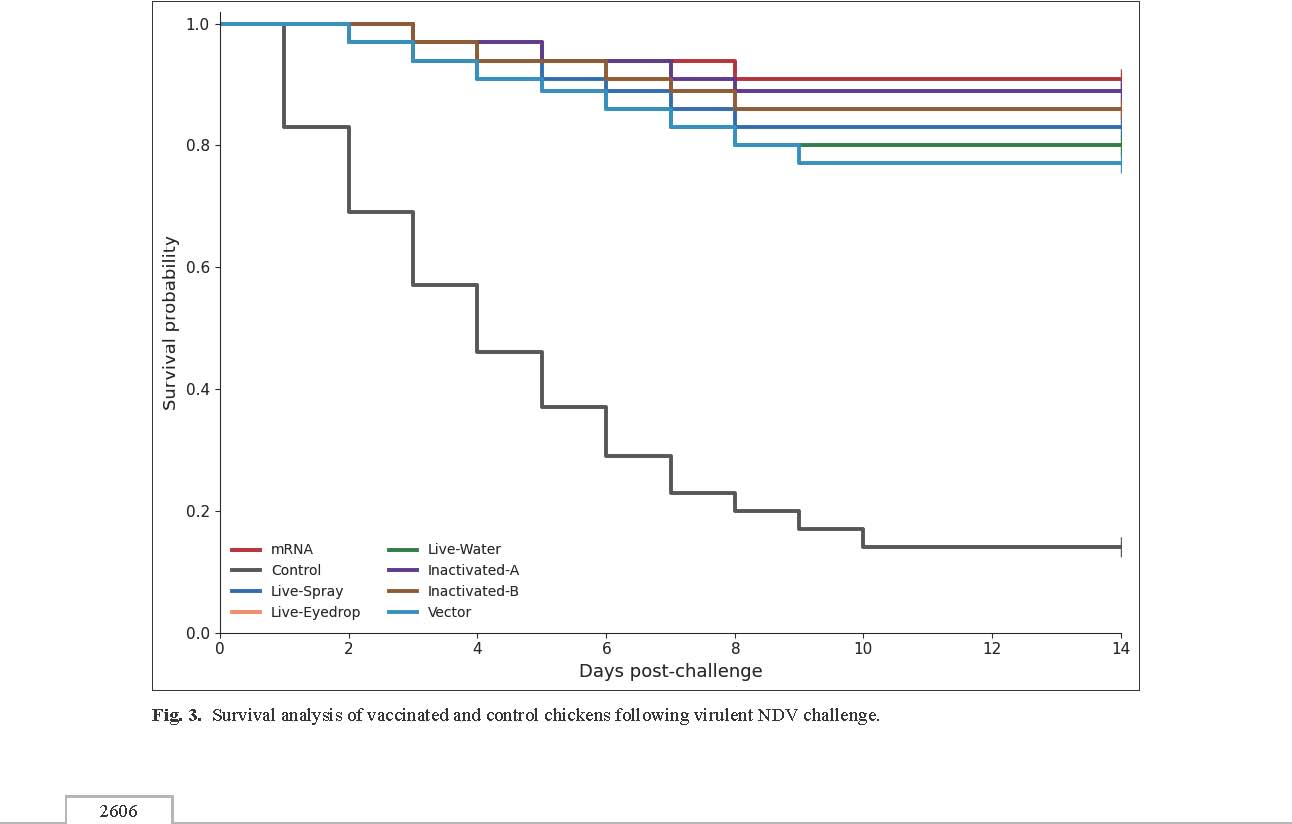
Fig. 2. Lymphocyte proliferation response. The bar graph shows lymphocyte proliferation expressed as the SI across the eight experimental groups. Clinical signs and morbidity following a virulent NDV challengeAfter the challenge with virulent NDV genotype VII, all birds in the unvaccinated control group had clinical signs compared with 22.9% (8/35) in the multi-epitope mRNA vaccine group. The commercial vaccine groups had clinical signs in 31.4%–45.7% of the birds, whereas 94.3% (33/35) of the birds in the unvaccinated control group exhibited clinical signs. The mean clinical score during the observation period was 3.8 ± 0.4 in the control group and 0.9 ± 0.2 in the mRNA-vaccinated groups, respectively (p < 0.001). The mean clinical scores in the vaccinated groups were 1.3 ± 0.3 to 1.8 ± 0.4. Survival rates and protective outcomesSurvival rates were analyzed over a 14-day post-challenge period. The unvaccinated control group had a survival rate of 14.3% (5/35). However, the multi-epitope mRNA vaccine group had a survival rate of 91.4% (32/35) (p < 0.001 vs. control). The commercial vaccine groups had survival rates of 71.4%–85.7%. The Kaplan–Meier survival curves show a clear separation between the control and vaccinated groups over the first 7 days after the challenge (Fig. 3).
Fig. 3. Survival analysis of vaccinated and control chickens following virulent NDV challenge. DiscussionThe present study showed that an mRNA vaccine targeting the conserved regions of the NDV F and the HN proteins is likely to induce strong and positive humoral and cellular immune responses. The NDV still poses a threat to the poultry industry despite the development of vaccines and efforts to control the disease. The development of an NDV vaccine that targets immune responses at the cellular and humoral levels will provide the industry with tools to reduce the incidence of this disease. The present study proved that the study by Raza et al. (2022)provided proof of concept that mRNA vaccines could be developed at the peptide level using computer elicitation. This study went further than Raza by using both F and HN components and integrating them into an mRNA platform. This study demonstrated that this integration provided positive results at the cellular and humoral immune levels. Response to genotype VII virus is a substantial step toward developing an NDV vaccine that targets cellular and humoral immune responses. The considerable humoral immune responses seen in participants vaccinated with mRNA showed considerable elevation in HI titers coupled with heightened levels of antibodies in ELISA, and they were found to be in alignment with the importance of both the HN and F proteins relating to the attainment of the virus and the merging of membranes. Haque et al. (2024)demonstrated an analogous elevation of antibody responses in the case of inactivated genotype VII.2 viruses from local mRNA isolates. Nevertheless, while inactivated vaccines operate on the principle of presenting all the viral antigens, in this case, the mRNA vaccine uniquely encodes the consensus sequences of the virus’s immunodominant epitopes. Such an approach may reduce antigenic noise and bias the immune response to the most important functional areas of the virus. Such an explanation may account for the elevated levels of antibodies, which were consistent and relatively elevated across all vaccinated participants. Vaccine strains and circulating NDV variants present a chronic obstacle to NDV control programs. Despite antigenic divergence, some studies have shown that classical vaccines offer partial protection against heterologous genotype VII strains (Hassanzadeh et al., 2024; Mahmood et al., 2024; Steensels et al., 2025). The present findings support and expand these studies by demonstrating how epitope-based mRNA vaccination can circumvent genotype mismatch by focusing on the highly conserved areas across genotype VII sublineages. This is especially important given the documented divergence and mutation in genotype VII viruses across multiple geographic regions, including Africa and the Middle East (Yadeta et al., 2024; Sabouri et al., 2025). In addition to humoral immunity, strong cell-mediated immune responses are also a significant benefit of the mRNA vaccine platform, which would be most relevant here. mRNA-vaccinated lymphocyte proliferation indices and IFN-γ–producing cell levels were significantly higher, indicating the potential of this platform to trigger T-cell-mediated immunity. This finding was essential because cellular immunity was critical for controlling the replication of the virus and, therefore, the amount of virus shed. This contrasts with several standard live and inactivated vaccines, which induce variable or suboptimal cellular immune responses despite adequate antibody responses (Ananda Kumar et al., 2023; Haque et al., 2024). It is also plausible that the more robust cellular responses in this case were related to the sophisticated intracellular antigen expression with MHC class I and II presentation that is characteristic of mRNA vaccines. The importance of a pattern of activated host immune genes after NDV infection was indicated in transcriptomic studies. For instance, Pandarangga et al. (2024) described immune transcriptional signatures in chickens that were vaccinated with genotype II compared to genotype VII vaccines, focusing on the differences in the activation of the adaptive and innate systems. Furthermore, in conjunction with the transcriptomic findings, the study findings demonstrated a balanced cellular and humoral response. They suggest that mRNA vaccines may induce a more balanced immune activation profile than other vaccine platforms. Protection outcomes following virulent challenge offer further rationalization for the multi-epitope mRNA vaccine immunological benefits. The mRNA-vaccinated group had a high survival rate, and clinical disease (respiratory distress (gasping), green watery diarrhea, and nervous signs such as torticollis or paralysis) was a good comparison for the protection provided by genotype-matched and genotype-mismatched vaccines for the recent challenge study (Hassanzadeh et al., 2024; Mahmood et al., 2024; Steensels et al., 2025). Of significance is that the protection was achieved without the need for whole virus, viral vectors, or nanoparticle encapsulation, which differentiates this approach from the recombinant or nanoparticle-based strategies of Sun et al. (2024)and Ananda Kumar et al. (2023). The rapid evolution and geographic spread of NDV genotype VII substantiate the vaccine design’s epidemiological relevance. Recent surveillance of molecules from Ethiopia and Iran indicate a more pronounced, evolving genetic drift and new sub-genotype formations that demonstrate greater divergence from classical vaccine lineages (Yadeta et al., 2024; Sabouri et al., 2025). Epitope-based mRNA, in this case, is a more appropriate design because these types of vaccines have the capability of rapid change as they can include newer conserved sequences that arouse as the virus evolves. The current results clearly showed that a multi-epitope mRNA vaccine focusing on conserved zones of the F and HN proteins can induce robust cellular and humoral responses and defend against NDV genotype VII infection. ConclusionIn addition to the successful in vivo performance of the vaccine, this study integrated the computational approach of vaccine design with immunoinformatics-epitope selection and modern nucleic acid vaccine platforms. The findings of this study warrant the advancement and refinement of the mRNA-based NDV vaccine as a paradigm-shifting mRNA vaccine for controlling Newcastle disease caused by the evolving and diversifying NDV genotypes. AcknowledgmentThe authors would like to express their gratitude to the College of Veterinary Medicine for providing laboratory facilities and technical support throughout the study. Conflict of interestThe authors declare that there are no conflicts of interest existed in this work. FundingThis research was self-funded by the authors with no external financial support. Authors’ contributions All authors have contributed to this study. Data availabilityData are available upon request from the corresponding author. ReferencesAnanda Kumar, B.S., Panickan, S., Bindu, S., Kumar, V., Ramakrishnan, S., Saxena, S., Shrivastava, S. and Dandapat, S. 2023. Immunogenicity and protective efficacy of an inactivated Newcastle disease virus vaccine encapsulated in poly-(lactic-co-glycolic acid) nanoparticles. Poultry Sci. 102(6), 102679; doi:10.1016/j.psj.2023.102679 Chowdhury, I.R., Viktorova, E., Samal, S.K. and Belov, G.A. 2024. The effect of 5′ and 3′ non-translated regions on the expression of a transgene from a Newcastle disease virus vector. Virus Res. 341, 199309; doi:10.1016/j.virusres.2024.199309 Haque, M.A., Haque, M.E., Parvin, M.K., Kamal, M.M., Islam, T.R., Sadekuzzaman, M., Islam, M.A., Khatun, M.M., Hossain, M.T., Uddin, M.A., Nahar, S.S., Khasruzzaman, A.K.M. and Islam, M.A. 2024. Determination of immunogenicity of an inactivated ND-vaccine developed experimentally with Newcastle disease virus (Genotype VII.2) local isolates of Bangladesh. Front. Immunol. 15, 1482314; doi:10.3389/fimmu.2024.1482314 Hassanzadeh, M., Abedi, M., Bashashati, M., Yousefi, A.R., Abdoshah, M. and Mirzaie, S. 2024. Evaluation of the Newcastle disease virus genotype VII-mismatched vaccines in SPF chickens: a challenge efficacy study. Vet. Anim. Sci. 24, 100348; doi:10.1016/j.vas.2024.100348 Ji, J., Dong, X., Liu, X., Ding, M., Lin, Y., Zhang, Y., Zhang, W., Lei, B., Yuan, W. and Zhao, K. 2025. Designing a novel multi-epitope trivalent vaccine against NDV, AIV and FAdV-4 based on immunoinformatics approaches. Microorganisms 13(12), 2744; doi:10.3390/microorganisms13122744 Kamran, Z., Waseem, M. and Asghar, M. 2026. In silico design of a matrix (M) protein-based vaccine candidate against Newcastle disease virus. Poultry Sci. 105(1), 106101; doi:10.1016/j.psj.2025.106101 Mahmood, S., Skinner, P., Warren, C.J., Mayers, J., James, J., Núñez, A., Lean, F.Z.X., Brookes, S.M., Brown, I.H., Banyard, A.C. and Ross, C.S. 2024. In vivo challenge studies on vaccinated chickens indicate a virus genotype mismatched vaccine still offers significant protection against NDV. Vaccine 42(3), 653–661; doi:10.1016/j.vaccine.2023.12.037 Mozafari, A., Amani, J., Shahsavandi, S. and Hatef Salmanian, A. 2022. A Novel multi-epitope edible vaccine candidate for newcastle disease virus: in silico approach. Iran. J. Biotechnol. 20(2), 3119. Pandarangga, P., Doan, P.T.K., Tearle, R., Low, W.Y., Ren, Y., Nguyen, H.T.H., Dharmayanti, N.I. and Hemmatzadeh, F. 2024. MRNA profiling and transcriptomics analysis of chickens received newcastle disease virus genotype II and genotype VII vaccines. Pathogens 13(8), 638; doi:10.3390/pathogens13080638 Randriamamisolonirina, N.T., Razafindrafara, M.S. and Maminiaina, O.F. 2025. Design of a multi-epitope vaccine against the glycoproteins of newcastle disease virus by using an immunoinformatics approach. ACS. Omega. 10(4), 4007–4018; doi:10.1021/acsomega.4c09890 Raza, A., Asif Rasheed, M., Raza, S., Tariq Navid, M., Afzal, A. and Jamil, F. 2022. Prediction and analysis of multi epitope based vaccine against Newcastle disease virus based on haemagglutinin neuraminidase protein. Saudi J. Biol. Sci. 29(4), 3006–3014; doi:10.1016/j.sjbs.2022.01.036 Sabouri, F., Kim, T.H., Abdoshah, M., Cho, A.Y., Fallah Mehrabadi, M.H., Lee, S.H., Song, C.S., Lee, D.H. and Bashashati, M. 2025. Evolution of Newcastle disease virus subgenotype VII.1.1 in Iran, 2017-2024. Avian Pathol. 55(2):171–82; doi: 10.1080/03079457.2025.2585135 Steensels, M., Soldan, C., Rauw, F., Roupie, V. and Lambrecht, B. 2025. Protective efficacy of classical vaccines and vaccination protocols against an exotic Newcastle disease virus genotype VII.2 in Belgian layer and broiler chickens. Poultry. Sci. 104(1), 104604; doi:10.1016/j.psj.2024.104604 Sun, W.Y., Cao, X.L., Wang, Y.X., Guo, X.C., Liu, J.M., Xue, Z.Q., Li, H.J., Wang, W., Zhang, T.T., Li, Q., Qin, R.H., Jin, Y.H., Li, Y.N. and Ren, G.P. 2024. Development and evaluation of a bivalent vaccine based on recombinant newcastle disease virus expressing infectious bursal disease virus VP2L-CH3-CH4 in SPF chickens. Vet. Microbiol. 288, 109950; doi:10.1016/j.vetmic.2023.109950 Tataje-Lavanda, L., Málaga, E., Verastegui, M., Mayta Huatuco, E., Icochea, E., Fernández-Díaz, M. and Zimic, M. 2023. Identification and evaluation in-vitro of conserved peptides with high affinity to MHC-I as potential protective epitopes for Newcastle disease virus vaccines. BMC. Vet. Res. 19(1), 196; doi:10.1186/s12917-023-03726-w Wu, Q., Liu, J., Cao, W., Li, J., Li, H., Jin, Y., Li, Z., Li, X., Sun, W., Bai, L., Shen, X., Guo, X. and Ren, G. 2025. GM-CSF-adjuvanted Newcastle disease virus-vectored bivalent vaccine elicits enhanced dual immunity against Newcastle disease and infectious bursal disease in chickens. Vet. Immunol. Immunopathol. 288, 111002; doi:10.1016/j.vetimm.2025.111002 Yadeta, W., Amosun, E., Mohammed, H., Woldemedhin, W., Sherefa, K., Legesse, A., Deresse, G., Birhanu, K., Abayneh, T., Getachew, B., Farnós, O., Kamen, A.A. and Gelaye, E. 2024. Isolation and genetic characterization of genotype VII velogenic pathotype newcastle disease virus from commercial chicken farms in Central Ethiopia, distinct from the local vaccine strains. Viruses 16(8), 1249; doi:10.3390/v16081249 Zeb, M.T., Dumont, E., Khan, M.T., Shehzadi, A. and Ahmad, I. 2024. Multi-epitopic peptide vaccine against newcastle disease virus: molecular dynamics simulation and experimental validation. Vaccines 12(11), 1250; doi:10.3390/vaccines12111250 | ||
| How to Cite this Article |
| Pubmed Style Shrshoh AA, Jasim NS. Different dose-based multi HN & F epitope mRNA vaccine against Newcastle disease. Open Vet. J.. 2026; 16(5): 2601-2609. doi:10.5455/OVJ.2026.v16.i5.3 Web Style Shrshoh AA, Jasim NS. Different dose-based multi HN & F epitope mRNA vaccine against Newcastle disease. https://www.openveterinaryjournal.com/?mno=308409 [Access: June 26, 2026]. doi:10.5455/OVJ.2026.v16.i5.3 AMA (American Medical Association) Style Shrshoh AA, Jasim NS. Different dose-based multi HN & F epitope mRNA vaccine against Newcastle disease. Open Vet. J.. 2026; 16(5): 2601-2609. doi:10.5455/OVJ.2026.v16.i5.3 Vancouver/ICMJE Style Shrshoh AA, Jasim NS. Different dose-based multi HN & F epitope mRNA vaccine against Newcastle disease. Open Vet. J.. (2026), [cited June 26, 2026]; 16(5): 2601-2609. doi:10.5455/OVJ.2026.v16.i5.3 Harvard Style Shrshoh, A. A. & Jasim, . N. S. (2026) Different dose-based multi HN & F epitope mRNA vaccine against Newcastle disease. Open Vet. J., 16 (5), 2601-2609. doi:10.5455/OVJ.2026.v16.i5.3 Turabian Style Shrshoh, Asma Abdlhusen, and Nafea Sabih Jasim. 2026. Different dose-based multi HN & F epitope mRNA vaccine against Newcastle disease. Open Veterinary Journal, 16 (5), 2601-2609. doi:10.5455/OVJ.2026.v16.i5.3 Chicago Style Shrshoh, Asma Abdlhusen, and Nafea Sabih Jasim. "Different dose-based multi HN & F epitope mRNA vaccine against Newcastle disease." Open Veterinary Journal 16 (2026), 2601-2609. doi:10.5455/OVJ.2026.v16.i5.3 MLA (The Modern Language Association) Style Shrshoh, Asma Abdlhusen, and Nafea Sabih Jasim. "Different dose-based multi HN & F epitope mRNA vaccine against Newcastle disease." Open Veterinary Journal 16.5 (2026), 2601-2609. Print. doi:10.5455/OVJ.2026.v16.i5.3 APA (American Psychological Association) Style Shrshoh, A. A. & Jasim, . N. S. (2026) Different dose-based multi HN & F epitope mRNA vaccine against Newcastle disease. Open Veterinary Journal, 16 (5), 2601-2609. doi:10.5455/OVJ.2026.v16.i5.3 |

