




| Review Article | ||
Open Vet. J.. 2026; 16(5): 2562-2580 Open Veterinary Journal, (2026), Vol. 16(5): 2562-2580 Review Article Current perspectives on avian tuberculosis in domestic and wild birds: Molecular diagnostics, genomic surveillance, and vaccination challengesYulianna Puspitasari1*, Aswin Rafif Khairullah2, Hartanto Mulyo Raharjo1, Wiwiek Tyasningsih1, Dea Anita Ariani Kurniasih3, Ikechukwu Benjamin Moses4, Bantari Wisynu Kusuma Wardhani5, Fitrine Ekawasti2 , Bima Putra Pratama6, Saifur Rehman7, Riza Zainuddin Ahmad2, Mutasem Abuzahra8, Fadhila Utari5, Syahputra Wibowo9, Ilma Fauziah Ma’ruf5 and Sri Suryatmiati Prihandani21Department of Veterinary Microbiology, Faculty of Veterinary Medicine, Universitas Airlangga, Surabaya, Indonesia 2Research Center for Veterinary Science, National Research and Innovation Agency (BRIN), Bogor, Indonesia 3Research Center for Public Health and Nutrition, National Research and Innovation Agency (BRIN), Bogor, Indonesia 4Department of Applied Microbiology, Faculty of Science, Ebonyi State University, Abakaliki, Nigeria 5Research Center for Pharmaceutical Ingredients and Traditional Medicine, National Research and Innovation Agency (BRIN), Bogor, Indonesia 6Research Center for Process Technology, National Research and Innovation Agency (BRIN), South Tangerang, Indonesia 7Department of Pathobiology, Faculty of Veterinary and Animal Sciences, Gomal University, Dera Ismail Khan, Pakistan 8Research Center for Animal Husbandry, National Research and Innovation Agency (BRIN), Bogor, Indonesia 9Eijkman Research Center for Molecular Biology, National Research and Innovation Agency (BRIN), Bogor, Indonesia *Corresponding Author: Yulianna Puspitasari. Department of Veterinary Microbiology, Faculty of Veterinary Medicine, Universitas Airlangga, Surabaya, Indonesia. Email: yulianna-puspitasari [at] fkh.unair.ac.id Submitted: 26/01/2026 Revised: 21/03/2026 Accepted: 01/04/2026 Published: 31/05/2026 © 2026 Open Veterinary Journal
ABSTRACTAvian tuberculosis (ATB) is a chronic infectious disease of poultry caused by Mycobacterium avium subsp. avium and is widespread in various regions of the world. This disease has a serious impact on poultry health and causes significant economic losses, primarily due to decreased body condition, reduced egg production, and increased mortality, particularly in young birds or individuals with compromised immune systems. Furthermore, ATB poses an opportunistic zoonotic risk, particularly for immunocompromised humans, highlighting its relevance within the One Health framework. This review aims to provide a comprehensive overview of ATB, covering etiology, epidemiology, pathogenesis, clinical symptoms, diagnostic methods, risk factors, economic implications, and prevention and control efforts. Unlike previous reviews, this study integrates recent findings on epidemiological patterns, zoonotic considerations, and current diagnostic and control approaches to provide an updated perspective on ATB in both domestic and wild bird populations. Infection transmission can occur through consumption of contaminated feed or water, exposure to aerosols, and direct or indirect contact with infected poultry or wild birds that act as reservoirs. A multimodal approach involving clinical evaluation, histopathology, acid-fast staining, bacterial culture, molecular methods, and serology is generally required to establish a diagnosis of ATB. Given the limited effectiveness of pharmacological therapies, control strategies are focused on biosecurity, improved environmental sanitation, isolation or culling of infected birds, and vaccine development. This review emphasizes the importance of implementing integrated control strategies to limit disease spread, maintain poultry health, reduce zoonotic risks, and minimize economic impacts. This review is expected to support the development of more effective surveillance and control strategies for ATB by synthesizing current knowledge and highlighting research gaps. Keywords: Avian tuberculosis, Mycobacterium avium, Disease, Poultry, Zoonosis. IntroductionAvian tuberculosis (ATB) is a chronic infectious disease of poultry caused by Mycobacterium avium subsp. avium (MAA), an acid-fast bacterium that can survive in the environment for long periods and cause systemic infections (Dhama et al., 2011). This disease has been reported to be widespread in domestic and wild birds, including chickens, turkeys, ducks, geese, pigeons, and various species of waterfowl (Abd El-Ghany, 2022). ATB is highly significant from an animal health perspective because it negatively impacts poultry productivity, characterized by weight loss, lethargy, reduced egg production, and increased mortality, particularly in young birds or individuals with compromised immune systems (Zhang et al., 2025). The chronic and slowly progressing nature of ATB makes early detection difficult, so cases are often only identified at an advanced stage, when granulomatous lesions have formed in internal organs such as the liver, spleen, bone marrow, and other lymphoid tissues (Batista et al., 2020). In addition to disrupting poultry health, ATB has significant economic consequences (Debelu et al., 2021). Chronic infection causes direct losses, including decreased production performance, increased mortality, and decreased quality of poultry products, such as eggs and meat (Liu et al., 2025). Furthermore, additional costs are required for implementing control measures, including waste and manure management, improved barn sanitation, isolation or culling of infected birds, and strengthening biosecurity systems to limit the spread of the disease (Butucel et al., 2022). The economic impact of ATB can accumulate significantly on an intensive poultry farm or commercial poultry industry scale, particularly in areas with high prevalence rates or suboptimal husbandry management practices (Mamo, 2020). ATB has also attracted attention due to its potential for zoonotic transmission, although human infections are relatively rare (Silva A del et al., 2009). Human cases are generally opportunistic and are more common in immunocompromised individuals, such as those with human immunodeficiency virus/acquired immunodeficiency syndrome or organ transplant recipients (Slany et al., 2016). Human exposure occurs primarily through direct contact with infected birds or through exposure to environments contaminated with the causative bacteria (Tsiouris et al., 2021). Therefore, ATB is included in the group of diseases relevant to the One Health approach, which emphasizes the interconnectedness of animal health, human health, and the environment in disease prevention and control strategies (Rabinowitz et al., 2013). The spread of ATB is influenced by various risk factors, including the age and immune status of poultry, population density, environmental conditions in cages, sanitation and biosecurity practices, and interactions with wild birds or other animals that act as reservoirs of the bacteria (Martin and Schimmel, 2000). Transmission occurs primarily through consumption of contaminated feed and water, exposure to aerosols, and direct and indirect contact with infected poultry (Levison, 2015). The slow-developing nature of the infection and the bacteria’s ability to persist in the environment make ATB control relatively difficult, necessitating an integrated approach encompassing prevention, surveillance, environmental management, and poultry population management (Kwaghe et al., 2015). This review aims to provide a comprehensive overview of ATB in poultry, covering etiology, pathogenesis, clinical presentation, pathological changes, host range, transmission mechanisms, risk factors, economic impact, diagnostic methods, and prevention and control efforts. A comprehensive understanding of these aspects is expected to serve as a reference for breeders, researchers, and veterinary practitioners in their efforts to detect, prevent, and effectively treat the disease, while simultaneously reducing the risk of interspecies bacterial spread and its resulting economic impact. EtiologyATB is caused by bacteria belonging to the Mycobacterium avium complex, with MAA as the primary etiologic agent in domestic poultry and wildlife (Bao et al., 2023). This bacterium is more accurately described as an acid-fast Bacillus characterized by a complex, lipid-rich cell wall containing mycolic acids than Gram-positive (Cook et al., 2009). The presence of these mycolic acids contributes to its strong resistance to staining, antimicrobial agents, and environmental stress (Sawaswong et al., 2025). MAA exhibits high resistance to various unfavorable environmental conditions, including exposure to disinfectants, extreme temperatures, and drought (Matos et al., 2025). MAA can survive for several months to more than 1 year in contaminated soil and poultry feces, particularly under moist and shaded conditions (Baratov and Mustafayev, 2024). These characteristics allow MAA to survive in soil, water, and poultry droppings for long periods, making the livestock environment a potential source of persistent infection (Whittington et al., 2004). In addition, the organism demonstrates relative resistance to commonly used disinfectants, such as quaternary ammonium compounds and some phenolic disinfectants, which further contributes to its environmental persistence (Falkinham, 2003). MAA is an obligate opportunistic pathogen (Agdestein et al., 2012). In poultry, infection generally occurs through the consumption of contaminated feed or water, although aerosol transmission can also occur, especially in high-density populations (Sattar et al., 2021). Mononuclear macrophages phagocytose MAA after it enters the host (Ignatov et al., 2012). However, bacteria survive by inhibiting the fusion process between phagosomes and lysosomes and evading cellular immune responses (Awuh and Flo, 2017). This mechanism allows MAA to replicate within phagocytic cells and subsequently trigger the formation of granulomatous lesions in various organs, particularly the liver, spleen, bone marrow, and lymphoid tissue (Daley, 2017). Several bacterial virulence factors have been identified, including genes involved in cell wall lipid biosynthesis, protective enzyme production against oxidative stress, and proteins capable of modulating the host immune response (Ssekitoleko et al., 2021). Genetic differences between strains also influence the level of virulence, the ability of the bacteria to survive in the environment, and the range of hosts they can infect (Shin and Shin, 2021). A comprehensive understanding of the etiology of the bacteria that cause ATB provides an essential basis for developing diagnostic methods, making disease control strategies, implementing biosecurity management, and exploring potential future vaccines. Host rangeATB has a broad host range, including domestic birds, wildlife, and several mammal species (Dhama et al., 2011). Infections have been reported in chickens, turkeys, ducks, geese, and various ornamental birds (Zhu et al., 2016). In addition, ATB has been documented in numerous wild bird species, including migratory birds, pigeons, and various waterbirds, which may act as natural reservoirs and contribute to environmental contamination and transmission to domestic poultry populations (Parvandar-Asadollahi et al., 2015). Figure 1 illustrates the overview of MAA and the main transmission pathways involved in ATB, highlighting the role of contaminated feces, feed, water, and environmental reservoirs in facilitating the spread of infection among susceptible bird populations.
Fig. 1. MAA and its primary transmission routes in ATB. MAA can also infect several mammalian species, particularly those that have close contact with birds or are exposed to contaminated environments, such as goats, cattle, pigs, and rodents (Shin and Shin, 2021). In mammals, infection is generally opportunistic and reported less frequently than in birds; however, it still has the potential to cause chronic disease characterized by the formation of granulomatous lesions similar to those found in birds (Daley, 2017). The specificity of the bacterial strain, the infectious dose, and the host’s immune system strongly influence the host range of ATB (Thom et al., 2008). Certain MAA strains exhibit host-adaptation patterns, where specific genotypes show higher pathogenicity and transmission efficiency in particular avian hosts, such as chickens or waterfowl, whereas other strains exhibit broader host plasticity and can infect multiple avian and mammalian species (Bao et al., 2023). Some MAA strains exhibit high virulence in specific poultry species, while others can infect a wider range of species (Schmidt et al., 2022). Interspecies interactions in both farm environments and natural ecosystems can increase the opportunity for cross-host transmission, making ATB a relevant disease within the One Health approach (Ferreira et al., 2024). EpidemiologyATB is a chronic infectious disease of birds that has a nearly worldwide distribution (Debelu et al., 2021). The disease is found in various types of domestic birds, such as chickens, turkeys, ducks, and geese, as well as in wild birds, which have the potential to act as reservoirs and transmission agents (Hernández, 2014). Environmental conditions, poultry management practices, and the level of interaction between domestic and wild bird populations influence the distribution of ATB (Kindu and Getaneh, 2016). ATB has been reported in almost every region of the world. In North America and Europe, the prevalence rates of bacteria in modern domestic poultry are relatively low, primarily because of stringent biosecurity practices, although the bacteria have been detected in wild bird populations and in the surrounding environment (Witte et al., 2008; Nesic et al., 2022). In contrast, higher incidence rates have been reported in Asia and Africa, particularly in areas with high livestock densities, inadequate sanitation, and limited animal health monitoring systems (Mayahi et al., 2013a; Zhang et al., 2025). However, variations in reported prevalence between regions may also reflect differences in diagnostic methods, surveillance intensity, and reporting systems used in epidemiological studies. Regions with more advanced laboratory capacity and systematic surveillance programs tend to detect and report cases more consistently, whereas areas with limited diagnostic infrastructure may experience underreporting (Motiwala et al., 2003). For example, several surveillance studies in Southeast Asian countries have isolated MAA from both domestic and wild birds, confirming the role of the environment as an important reservoir in the transmission of infection (Lâm et al., 2025). Geographical conditions and ecological factors also play a role in determining ATB regional distribution patterns (Li et al., 2022). Areas with high humidity and moderate to low temperatures tend to favor the survival of bacteria in the environment, increasing the likelihood of infection in poultry (Baratov and Mustafayev, 2024). The movement and migration of wild birds between countries also contribute to the spread of the disease, making ATB a transboundary animal health issue (Olowu et al., 2026). Various epidemiological surveys report wide variations in prevalence, with incidence rates of less than 1% in commercial poultry operations with strict biosecurity practices but exceeding 20% in traditional or backyard production systems with limited hygiene and disease monitoring (Debelu et al., 2021). These estimates may refer to either bird-level prevalence (the proportion of infected individuals within a flock) or flock-level prevalence (the proportion of affected flocks within a surveyed population), depending on the study design and surveillance method used (Verdugo et al., 2014). Additionally, prevalence estimates may vary depending on the diagnostic approach applied, such as bacteriological culture, molecular detection, or postmortem identification of characteristic lesions (Blanco Vázquez et al., 2020). The global and regional distribution patterns of ATB underscore the importance of ongoing surveillance, adequate biosecurity practices, and the integration of a One Health approach (Pereira et al., 2020). This is because ATB not only impacts poultry health and livestock productivity but also poses potential zoonotic risks, particularly to individuals with compromised immune systems (Libera et al., 2022). A comprehensive understanding of this disease’s geographic distribution is crucial for designing effective control strategies and preventing its spread. PathogenesisATB pathogenesis is a complex process determined by the dynamic interaction between MAA and the host bird immune system (Miggiano et al., 2020). Infection generally begins when birds ingest contaminated feed or water or inhale bacteria-containing aerosols (Levison, 2015). MAA is then phagocytosed by mononuclear macrophages found in the mucosa of the digestive and respiratory tracts after entering the body (Maina, 2023). MAA can survive inside macrophage phagosomes by inhibiting the fusion process between phagosomes and lysosomes and suppressing macrophage activation, allowing the bacteria to replicate without being eliminated by the host’s cellular defense mechanisms (Wang and Lee, 2025). This intracellular survival is partly mediated by specific virulence mechanisms, including ESAT-6 secretion system (type VII) secretion systems that facilitate the export of effector proteins involved in immune modulation and intracellular persistence (Roy et al., 2020). The presence of mycolic acids in the bacterial cell wall contributes to immune evasion by limiting phagolysosomal maturation and protecting the bacterium from host antimicrobial responses (Wang et al., 2023). Bacteria inside macrophages trigger a typical host immune response, characterized by the formation of granulomas (Abukhalid et al., 2021). These granulomatous structures are composed of infected macrophages, multinucleated giant cells, lymphocytes, and fibrotic tissue in some cases (Joseph et al., 2023). Granulomas frequently develop central caseous necrosis in avian species; however, the lesions tend to be more compact and less extensively fibrotic compared with those observed in mammalian tuberculosis (Flórido and Appelberg, 2004). In birds, fibrosis is often less pronounced and may appear as thin fibrous capsules surrounding granulomatous nodules rather than the dense fibrotic structures typically seen in mammals (Dhama et al., 2011). Although granulomas play a role in limiting bacterial spread, they can also cause tissue damage and impaired organ function, particularly in the liver, spleen, bone marrow, and other lymphoid tissues (Acharya et al., 2018). The pathogenesis of MAA is influenced by various virulence factors, including cell wall lipid biosynthesis, which increases bacterial resistance to oxidative stress, protein production that suppresses the host immune response, and the ability of the bacteria to modulate phagocytic cell metabolism to support intracellular growth (Agdestein et al., 2014). Additional factors contributing to bacterial persistence include enzymes that protect against oxidative stress, such as catalase-peroxidase and superoxide dismutase, which neutralize reactive oxygen species produced by host immune cells (Forrellad et al., 2013). Furthermore, genetic differences between strains contribute to variations in virulence levels, host preferences, and the capacity of bacteria to survive in the environment (Kannan et al., 2019). In addition to bacterial factors, the immune status and general health of the birds also play a significant role in determining the infection severity (Matos et al., 2025). Young birds and individuals experiencing stress or malnutrition are more susceptible to developing more severe lesions (Roller et al., 2020). The disease typically progresses slowly over weeks to months, beginning with early macrophage infection and microscopic granuloma formation, followed by visible nodular lesions in internal organs (Shivaprasad and Palmieri, 2012). Progressive granuloma accumulation can lead to tissue damage, impaired organ function, reduced production performance, and death in chronic cases (Koets et al., 2015). Multiple granulomas may coalesce in advanced stages, resulting in extensive organ involvement and the characteristic chronic clinical presentation of ATB (Carabalí-Isajar et al., 2023). Immune responseIn birds, ATB infection elicits a complex immune response through the involvement of both the innate and adaptive immune systems (Kaiser, 2010). After the bacteria enter the body through the digestive or respiratory tract, macrophages and dendritic cells act as the first line of defense (To et al., 2020). MAA is then phagocytosed by macrophages, where it can persist intracellularly and modulate host immune signaling pathways, thereby reducing the effectiveness of Mycobacterium abscessus complex (Gomes et al., 1999). The adaptive immune response is then induced through antigen presentation by macrophages and dendritic cells to cluster of differentiation 4 positive (CD4+) T lymphocytes and cluster of differentiation 8 positive (CD8+) T lymphocytes (Basler et al., 2013). The T helper 1 (Th1) response is characterized by the secretion of interferon-gamma (IFN-γ) and tumor necrosis factor-alpha, which play a crucial role in activating macrophages and facilitating granuloma formation (Mendoza-Trujillo et al., 2025). Granulomas function as a defense mechanism by limiting the spread of bacteria by grouping infected cells in an organized structure composed of macrophages, multinucleated giant cells, lymphocytes, and fibrotic tissue (Lyu et al., 2024). However, MAA possesses various mechanisms to evade the host immune response, including the ability to modulate host cell apoptosis, increase resistance to oxidative stress, and undergo surface antigen variation (Kim et al., 2021). These mechanisms allow bacteria to persist within granulomas for extended periods (Danelishvili et al., 2020). The immune response is often suboptimal in young birds or individuals with compromised immune systems, allowing granulomas to develop into large lesions and cause progressive tissue damage to vital organs, such as the liver, spleen, and bone marrow (Tsiouris et al., 2021). In addition, the formation of antibodies against MAA antigens can identify humoral immune responses; however, these antibodies are generally ineffective in eradicating infection (Achkar and Casadevall, 2013). Despite their limited role in bacterial clearance, antibody responses are still important for diagnostic purposes, as serological tests can detect exposure to MAA in infected flocks. Higher antibody titers have been associated with advanced or chronic stages of infection in some cases, although this relationship is not always consistent due to variability in host immune responses and infection stages (Roque et al., 2023). Serological assays in birds may have limitations related to cross-reactivity with other environmental mycobacteria and variability in antibody production among individuals, which can affect the sensitivity and specificity of these tests (Turenne et al., 2007). Therefore, ATB control relies more on cellular immunity mechanisms (Dhama et al., 2011). A thorough understanding of these immunological processes is crucial for developing immunology-based diagnostic methods, exploring potential vaccines, and implementing biosecurity-based disease management strategies, particularly in high-density poultry populations with increased susceptibility. PathologyATB causes characteristic pathological changes that reflect a complex interaction between the causative agent and the avian immune response (Kwaghe et al., 2015). After the bacteria enter the body through the digestive or respiratory tract, MAA replicates within macrophages, resulting in the development of granulomatous lesions in multiple internal organs, particularly the liver, spleen, bone marrow, and other lymphoid tissues (Li et al., 2010). These lesions represent the pathological consequence of the host immune response attempting to contain the infection (Schinköthe et al., 2016). Although they play a role in limiting bacterial spread, extensive granuloma formation can lead to tissue damage and impaired organ function (Khairullah et al., 2024). Macroscopically, ATB lesions generally appear as small, yellowish-brown to yellowish-white nodules on organ surfaces, which can be multifocal or widespread (Dvorska et al., 2007). These nodules often undergo central necrosis and calcification in the chronic stage, resulting in a tissue appearance resembling a caseous mass (Flórido and Appelberg, 2004). Granulomas can cause organ enlargement, particularly in the spleen and liver, which appear hypertrophic on postmortem examination (Acharya et al., 2018). Microscopically, the lesions typically show organized granulomatous inflammation with necrotic centers, cellular debris accumulation, and varying degrees of mineralization, which are characteristic features of advanced infections (Severova et al., 2025). However, similar granulomatous or nodular lesions may also occur in other avian diseases, making differential diagnosis essential. Conditions that may mimic ATB lesions include neoplastic diseases such as avian leukosis and Marek’s disease, as well as fungal granulomatous infections such as aspergillosis (Flórido and Appelberg, 2004). Therefore, additional diagnostic methods, including acid-fast staining, bacterial culture, or molecular detection techniques, should be used to confirm the presence of MAA (Mayahi et al., 2013b). In addition to attacking internal organs, MAA can also infect subcutaneous tissue and the gastrointestinal tract, triggering chronic inflammatory cell infiltration and intestinal mucosa granuloma formation (Lyu et al., 2024). The development of these pathological changes is strongly influenced by the bird’s immune status, age, and infectious dose, with young birds or individuals with compromised immune status generally showing more extensive and rapidly progressing lesions (Mariappan et al., 2023). Clinical manifestationsATB generally develops slowly with a chronic course and presents with a variety of clinical manifestations, depending on the bird species, age, immune status, and extent of exposure (Kaevska et al., 2010). In the early stages, infection in birds is often subclinical, so the presence of the disease is often missed without laboratory testing or postmortem evaluation (Ncube et al., 2022). In cases with clear clinical manifestations, birds may experience weight loss, lethargy, decreased appetite, and decreased egg production in laying hens (Stepień-Pyśniak et al., 2016). Some individuals appear to have poor body condition or become emaciated despite adequate feed intake (Gill et al., 2011). Gastrointestinal disturbances, such as diarrhea or paler stool color, may occur as a result of gastrointestinal tract lesions (Komatsu et al., 2017). In some species, particularly young chickens and turkeys, severe infections can lead to sudden death. Figure 2 illustrates the visible clinical manifestations associated with ATB in infected poultry, highlighting common signs such as progressive weight loss, lethargy, ruffled feathers, reduced egg production, and intermittent diarrhea resulting from infection by MAA.
Fig. 2. Visible clinical manifestations of ATB in infected poultry. Physical examination can sometimes identify enlarged internal organs such as the liver, spleen, and lymphoid tissue, reflecting the accumulation of granulomatous lesions in these organs (Darya et al., 2025). However, external clinical signs are often nonspecific and can mimic a variety of other chronic avian diseases, including long-standing bacterial or parasitic infections (Slany et al., 2016). Therefore, establishing a diagnosis based on clinical symptoms alone is generally inadequate, requiring additional examinations, such as laboratory analysis, histopathological evaluation, or molecular techniques, to confirm the diagnosis (Song et al., 2026). The age and immune status of the birds also influence the clinical manifestations of ATB (Kwaghe et al., 2015). Young birds and individuals with compromised immune status generally exhibit more severe and progressive symptoms, whereas adults often experience infections that remain asymptomatic for long periods (Debelu et al., 2021). The chronic nature of the disease and its nonspecific clinical signs make ATB a major challenge for early detection and control in poultry systems (Dhama et al., 2011). DiagnosisATB diagnosis is a crucial component of disease control efforts, given that the infection is chronic, progresses slowly, and often lacks typical clinical signs. Figure 3 illustrates the multimodal diagnostic challenges and laboratory confirmation strategies for ATB, highlighting the integration of clinical evaluation, postmortem lesion identification, histopathological examination, bacteriological culture, and molecular methods to accurately detect infection caused by MAA in poultry populations. Infected birds may exhibit symptoms such as lethargy, weight loss, reduced egg production, or emaciation; however, these manifestations are also common in many other chronic avian diseases (Srivastava et al., 2017). However, laboratory confirmation is essential for accurate diagnosis because these manifestations are not specific indicators of ATB (Siddiqui et al., 2025).
Fig. 3. Multimodal diagnostic challenges and laboratory confirmation strategies for acute tuberculosis. One of the initial stages in identifying ATB is postmortem examination, where characteristic granulomatous lesions can be found in organs such as the liver, spleen, bone marrow, and lymphoid tissue (Moravkova et al., 2011). Although the presence of granulomas is an important indicator, laboratory confirmation is still necessary because these lesions are not completely specific (Tsiouris et al., 2021). Histopathological analysis using acid-fast staining allows visualization of the bacteria within the tissue, while isolation of the bacteria through culture from internal organs provides the most definitive diagnostic evidence and is considered the gold standard for confirming infection (Lari Baghal et al., 2024). Nevertheless, bacteriological culture has several limitations, including the slow growth rate of MAA, which may require several weeks to obtain visible colonies, the need for specialized mycobacterial culture media, and the risk of contamination by other environmental microorganisms (Park et al., 2024). These factors can delay diagnostic confirmation and complicate timely decision-making in disease control programs. In addition to conventional approaches, molecular methods—particularly polymerase chain reaction (PCR)—have emerged as rapid and highly sensitive diagnostic tools for detecting MAA deoxyribonucleic acid (DNA), including in subclinical cases or conditions with low bacterial loads (Singh and Kashyap, 2012). Serological and tuberculin skin tests can also be used as screening methods in poultry populations, although their sensitivity and specificity are highly influenced by immune status, age, and environmental conditions (Angkwanish et al., 2021). From a practical perspective, each diagnostic method has specific advantages and limitations. Clinical observation and postmortem examination are useful for initial suspicion and field-level screening but lack specificity (Woodbury et al., 2008). Histopathology and acid-fast staining provide supportive evidence of characteristic granulomatous inflammation and acid-fast bacilli in tissues (Daley, 2017). Bacteriological culture remains the gold standard for definitive confirmation, although the process is time-consuming and technically demanding (Dane et al., 2022). In contrast, molecular techniques such as PCR offer faster detection with high sensitivity and specificity, making them particularly valuable for early diagnosis and surveillance programs (Sevilla et al., 2015). An effective diagnostic approach for ATB is generally multimodal, integrating clinical assessment, postmortem examination, histopathological analysis, culture isolation, and molecular techniques (Srivastava et al., 2017). This strategy allows for earlier case identification, differentiation of ATB from other diseases with similar clinical manifestations, and implementation of more appropriate and efficient control measures in both domestic and wild bird populations (Zhang et al., 2025). Differential diagnosisATB often exhibits clinical features and internal pathological changes that mimic various chronic diseases in poultry (Algammal et al., 2021). Because the disease frequently develops gradually and may resemble several other chronic infections, careful pathological and laboratory evaluation is required to distinguish ATB from other conditions (Abd El-Ghany, 2022). In veterinary practice, several other diseases need to be considered as differential diagnoses (Dhama et al., 2011). Salmonella pullorum and Salmonella gallinarum infection, which cause pullorum disease and Fowl typhoid, respectively, can cause lethargy, diarrhea, and increased mortality, especially in young chickens (Kang et al., 2022). Although the enlargement of internal organs such as the liver and spleen is common, the characteristic granulomatous lesions of ATB are generally absent (Haque et al., 2016). Furthermore, lymphoid leukosis—a disease caused by a retrovirus that attacks lymphocytes—can cause lymphoid tissue enlargement and nodule formation in the liver, but without granuloma formation. Histopathological examination or viral detection by PCR is required for diagnosis (Elmeligy et al., 2024). In addition, systemic fungal infections, such as those caused by Aspergillus or Candida, can produce internal granulomatous lesions that resemble the granulomas of ATB (Kubota et al., 2023). The use of special stains, such as Periodic acid–Schiff or Grocott methenamine silver, helps differentiate fungal-infected tissue from bacterial granulomas (Guarner and Brandt, 2011). Infections with other bacteria, such as Listeria monocytogenes, Staphylococcus spp., or Clostridium spp., can also form granulomas. However, bacterial culture isolation and acid-fast staining remain the primary methods for confirming the diagnosis of ATB (Caulfield and Wengenack, 2016). Furthermore, chronic conditions resulting from malnutrition or vitamin deficiency can produce clinical symptoms and organ damage similar to those of ATB infection, even though typical granulomas are not formed (Lyu et al., 2024). Therefore, ATB differentiation from these conditions relies on a combination of pathological examination and specific laboratory confirmation methods, including acid-fast staining, bacterial culture, and molecular detection techniques such as PCR (Tell et al., 2003). A thorough understanding of this differential diagnosis is crucial for avoiding misdiagnosis, implementing appropriate control strategies, and minimizing economic losses in poultry systems. TransmissionThe long-term survival of MAA in the environment influences the spread of ATB, allowing the disease to be transmitted through various routes (Slany et al., 2016). Infection in poultry generally occurs through the consumption of feed or water contaminated by feces, dust, or environmental materials containing MAA (Abd El-Ghany, 2022). In addition to the oral route, inhalation of aerosol particles containing the bacteria can also be a transmission mechanism, especially in high-density poultry populations, where dust and droplets are easily inhaled (Debelu et al., 2021). Infected poultry can excrete MAA through feces and respiratory secretions and may also contaminate eggs during the laying process (Yavuz et al., 2021). Feces containing these bacteria can potentially contaminate the cage environment, feed, drinking water, and equipment, thereby becoming a source of secondary infection for other birds (Moravkova et al., 2011). Distinguishing between true transovarial transmission and external eggshell contamination is important for egg-associated transmission (Nielsen et al., 2004). Current evidence suggests that true transovarial transmission—where the bacteria infect the developing embryo through the reproductive tract—occurs rarely (Bielanski et al., 2006). MAA contaminates the external surface of eggs through contact with infected feces or contaminated nesting materials, which can subsequently expose chicks during hatching if proper hygiene is not maintained (Shitaye et al., 2008). Lax biosecurity practices, inadequate cage sanitation, and direct contact between domestic poultry and wild birds, which act as natural reservoirs for the bacteria, further exacerbate the spread of the disease (Kwaghe et al., 2015). Some mammals exposed to contaminated environments can also be infected with MAA, although cases in mammals are relatively rare and generally opportunistic (Reil et al., 2024). Migratory wild birds play a significant role in the spread of disease across regions, as they can carry the bacteria from one location to another, increasing the risk of transmission between domestic and wild bird populations (Olowu et al., 2026). Because environmental contamination is the dominant transmission pathway, effective control strategies primarily focus on sanitation, proper waste management, egg-handling hygiene, and strict hatchery biosecurity to prevent cross-contamination during incubation and hatching (Dvorska et al., 2007). Understanding ATB transmission pathways is crucial for designing biosecurity-based control strategies, including waste management, pen sanitation, limiting contact with wild birds, and preventing feed and water contamination. Implementing these strategies is key to suppressing disease spread while minimizing economic losses in the poultry industry. Risk factorsThe spread of ATB is influenced by host conditions, the environment, and farm management practices (Kwaghe et al., 2015). Understanding these determinants is essential for identifying the conditions that facilitate the persistence and circulation of MAA within poultry production systems (Cromie et al., 1991). From a host perspective, the age and immune status of birds are the main factors influencing susceptibility to ATB (Hasan et al., 2016). Young birds may show increased vulnerability to infection due to the incomplete development of their immune defenses, and infection in these individuals may progress more rapidly than in adult birds (Roller et al., 2020). Sensitivity to MAA can also differ between species, resulting in varying susceptibility levels (Fernandez-Pittol et al., 2022). Physiological stress and poor nutritional status can reduce the overall resilience of birds, indirectly increasing the likelihood of infection and disease progression (Matos et al., 2025). The farm environment is also a significant risk factor (Villaamil et al., 2021). Conditions that favor the accumulation and long-term survival of the bacteria—such as damp litter, poor hygiene of housing facilities, and inadequate waste management—can increase the probability of exposure within flocks (Kriz et al., 2013; Lechevallier et al., 2024). Factors related to farm management also influence the spread of ATB (Kwaghe et al., 2024). Insufficient cleaning and disinfection routines, inadequate manure disposal, lack of quarantine procedures for newly introduced birds, and poor flock health monitoring can facilitate the maintenance of infection sources within poultry operations (Corn et al., 2005). These management-related factors may enable the pathogen to persist in production systems for extended periods if control measures are not consistently applied (Coleman et al., 2022). The interaction between domestic poultry and surrounding wildlife populations may contribute to environmental contamination and indirect exposure to MAA in certain production settings (Shitaye et al., 2008; Soler et al., 2009). Consequently, comprehensive risk assessment and targeted management interventions are essential for reducing the likelihood of infection and supporting sustainable disease control programs (Garvey and M, 2020). Economic impactATB significantly impacts the poultry industry, particularly in areas with poor biosecurity and suboptimal management practices (Dhama et al., 2011). Because the disease progresses chronically, early detection is often difficult, resulting in prolonged productivity losses in domestic poultry populations (Liu et al., 2025). The economic consequences of ATB can generally be categorized into direct production losses and indirect costs associated with disease control and long-term management (Lekko et al., 2021). Direct losses primarily include reduced productivity, impaired growth performance, and mortality within infected flocks (Dhama et al., 2011). One of the main impacts of ATB is decreased egg production and poultry growth (Debelu et al., 2021). Chronic infection may reduce egg production by approximately 10%–20% in affected laying flocks, depending on disease severity and flock management conditions (Kaboudi et al., 2017). Infected birds often exhibit lethargy, reduced appetite, and weight loss, resulting in decreased feed conversion efficiency and increased production costs (Tell et al., 2001). Growth rate impairment of up to 15%–30% due to chronic wasting and reduced nutrient utilization has also been reported in severely affected birds (Riber and Wurtz, 2024). In laying hens, chronic infection can reduce both egg quantity and quality, directly impacting the farmer’s economic losses (Roberts et al., 2011). Furthermore, ATB can cause gradual mortality in poultry, particularly in young birds or individuals with weakened immune systems (Kaboudi et al., 2017). Mortality rates in infected flocks may vary widely but have been reported to reach 5%–15% in poorly managed systems or in advanced stages of infection (Salamatian et al., 2020). This mortality reduces the number of livestock available for production, impacting farmer incomes and the entire poultry supply chain (Grace et al., 2024). Furthermore, the chronic nature of the disease forces farmers to incur additional costs for feed, care, and environmental management, increasing operational costs (Debelu et al., 2021). Long-term disease control measures and regulatory consequences cause indirect economic losses. The economic impact of ATB extends beyond production losses (Silva et al., 2021). Infection also incurs additional costs related to control and biosecurity measures, such as isolating infected birds, disinfecting cages, and managing contaminated waste (Abd El-Ghany, 2022). Additional expenditures may include surveillance programs, diagnostic testing, flock monitoring, and stricter biosecurity protocols to prevent infection recurrence (Mehmedi et al., 2025). In regions or countries with high prevalence, the disease can also impact the trade in poultry and poultry products due to the risk of cross-border transmission and strict animal health regulations (Mai et al., 2024). Consequently, ATB can impose long-term economic burdens on both individual farmers and the poultry industry as a whole through decreased productivity, increased management costs, and potential trade limitations (Garvey and M, 2020). Zoonotic potentialAlthough ATB has zoonotic potential, human infections are rare. The bacterium MAA, which belongs to the MAC group, can infect a wide range of hosts, including birds, mammals, and humans, especially those with weakened immune systems (Slany et al., 2016). Human exposure generally occurs through direct contact with infected birds, cage environments contaminated with feces or dust, or consumption of unhygienically prepared poultry products (Debelu et al., 2021). Human MAA infections are generally opportunistic, occurring in immunocompromised individuals, such as those with HIV/AIDS, organ transplant recipients, or those with other chronic illnesses (To et al., 2020). Clinical manifestations can include respiratory tract infections, the formation of granulomatous lesions in the lungs, or systemic infections in severely immunocompromised individuals (Carabalí-Isajar et al., 2023). Although human-to-human transmission is extremely rare, the presence of MAA in crowded poultry environments increases the risk of exposure for farm workers, veterinary personnel, and populations with frequent contact with poultry (Reil et al., 2024). The zoonotic risk of ATB is influenced by several factors, including MAA prevalence in poultry, individual immune status, environmental sanitation conditions, and biosecurity practice implementation (Reza et al., 2015). Subclinically infected poultry can serve as a reservoir of the bacteria without showing obvious symptoms, increasing the likelihood of human exposure (Knific et al., 2022). Furthermore, wild birds and other animals that act as natural reservoirs can spread MAA across locations, expanding the scope of the zoonotic risk (Lekko et al., 2021). TreatmentThe pharmacological treatment of ATB in poultry is very limited in effectiveness (Buur and Saggese, 2012). The infection progresses slowly, MAA can persist within macrophages for long periods, and granuloma formation prevents drug penetration (Charrier et al., 2025). Therefore, antibiotic therapy is rarely successful and routinely used in commercial poultry practices (Salamatian et al., 2020). Several antibiotics, such as rifampin, isoniazid, and ethambutol, commonly used to treat MAA infections in mammals, have been experimentally tested in poultry (Saggese et al., 2014). Although these drugs can reduce bacterial numbers, their application faces several challenges (Dao et al., 2025). The long treatment duration, often spanning several weeks to months, makes it difficult to implement in large poultry populations (Harris and Barletta, 2001). Furthermore, long-term antibiotic use risks the development of bacterial resistance and drug residues in poultry products, such as meat and eggs, which raises food safety concerns (Nhung et al., 2017). Due to the limitations of pharmacological treatments, strategies for managing ATB in poultry focus more on prevention and environmental control than on individual therapy (Abd El-Ghany, 2022). Infected birds are generally isolated or culled to prevent the spread of the disease, while barn sanitation, waste management, and strict biosecurity practices are key control measures (Kwaghe et al., 2024). Controlling contact with wild birds or other animals that act as bacterial reservoirs is also a crucial component of disease management strategies (Kobuszewska and Wysok, 2024). Several experimental studies have evaluated combinations of antibiotic therapy with immunomodulators or vaccines to enhance the immune response and suppress bacterial numbers, but their application in commercial farms remains limited (Park and Yoo, 2016; Rais et al., 2023). Therefore, pharmacological treatment of ATB in poultry is largely experimental and not a primary strategy (Ledwoń et al., 2015). More effective approaches continue to focus on prevention, environmental control, and biosecurity, which reduce disease spread and minimize economic losses (Hernández-Agudelo et al., 2025). VaccinationVaccination is a potential strategy for the prevention and control of ATB, a chronic disease caused by MAA in poultry (Sattar et al., 2021). Given the slow progression of infection and the long-term persistence of the bacteria in the environment, vaccination plays a role in enhancing the cellular immune response in poultry against MAA, thereby reducing disease incidence, granulomatous lesion formation, and the risk of transmission within the population (Park and Yoo, 2016). To date, the use of ATB vaccines in domestic poultry remains limited, and live, attenuated vaccines are mostly used, similar to the principle of the BCG vaccine in humans (de Lisle et al., 2005). These vaccines are designed to stimulate a Th1 immune response, specifically through the production of IFN-γ, macrophage activation, and granuloma formation, which effectively suppress bacterial replication (Park et al., 2022). Although vaccination can reduce the severity of lesions, reduce the number of bacteria released into the environment, and slow the spread of the disease, it does not always prevent infection completely (Orujyan et al., 2022). Vaccine development for ATB faces several challenges. Genetic variability between MAA strains can impact vaccine effectiveness, while the bacteria’s ability to survive within macrophages limits the achievement of an optimal immune response (Barletta et al., 2023). Furthermore, vaccination can affect the interpretation of tuberculin skin tests and serological tests, requiring the tailoring of diagnostic strategies to vaccination programs (Gupta et al., 2023). This issue is closely related to the differentiating infected from vaccinated animals (DIVA) challenge, where conventional diagnostic methods may be used to distinguish vaccinated birds’ immune responses from those generated by natural infection (Koo et al., 2004). The absence of reliable DIVA-compatible vaccines complicates surveillance programs and may limit the large-scale implementation of vaccination strategies in poultry populations (Tsiouris et al., 2021). Other factors that influence vaccine response include the age of the birds, their immune status, and environmental conditions (Howard et al., 2002). Several experimental studies have also explored the use of subunit and DNA vaccines to improve safety and efficacy (Rathnaiah et al., 2017; Boroojeni et al., 2025). These next-generation vaccine approaches are particularly promising for addressing field limitations because they may allow the development of DIVA-compatible vaccines where specific antigens differ from those targeted by diagnostic tests, enabling accurate differentiation between vaccinated and naturally infected birds (Park and Yoo, 2016). These approaches target specific MAA antigens, thereby stimulating a more targeted immune response, minimizing the risk of side effects, and facilitating the differentiation between vaccinated and naturally infected birds (Matos et al., 2025). ControlControlling ATB is a crucial component of poultry health management, given that the disease is chronic, progresses slowly, and is difficult to treat pharmacologically (Debelu et al., 2021). Control strategies emphasize prevention, environmental management, biosecurity, and poultry population management, as antibiotic use is rarely effective and can potentially lead to drug residues and bacterial resistance (Clark et al., 2012). The primary strategy for controlling ATB involves identifying and eliminating infected birds (Abd El-Ghany, 2022). Systematic culling of birds diagnosed with ATB or showing granulomatous lesions is crucial in preventing the spread of the bacteria to healthy populations (Zhang et al., 2025). Routine monitoring and surveillance must account for the interaction between vaccination and diagnostic tests, particularly in flocks that may have received non-DIVA vaccines, to avoid misclassification of vaccinated birds as infected (Chui et al., 2004). Clinical examination, histopathology, tuberculin skin tests, and molecular methods such as PCR are essential for detecting subclinical cases or contaminated populations (Borham et al., 2022). Environmental management plays a crucial role in the control of ATB (Kwaghe et al., 2015). Maintaining clean pens, properly managing manure, drying out the pen area, and controlling humidity and dust can reduce bacterial concentrations in the environment (Grewal et al., 2006). Equipment, feed, and drinking water should also be kept clean and protected from contamination by wild birds or other animals that act as natural reservoirs of ATB (Dvorska et al., 2007). Biosecurity implementation includes isolating new birds before they are introduced to the main population, regulating human and vehicle access to the enclosure, and limiting interaction with wild birds and other animals (Ngom et al., 2024). Specific structural modifications are recommended to reduce contact with wildlife reservoirs in addition to general biosecurity practices (Lombard and Garry, 2025). These include installing high-density netting or wire mesh around poultry houses to prevent the entry of wild birds, using covered or indoor feeders to minimize feed contamination, and protecting drinking water sources through enclosed water systems or overhead covers to prevent contamination from wild waterfowl droppings (Reil et al., 2024). The regular inspection and maintenance of these protective structures are also essential to ensure their effectiveness in preventing wildlife intrusion. These measures are essential for preventing the introduction of external bacteria and reducing the risk of transmission among birds within the population (Butucel et al., 2022). Research challenges and prospectsATB research faces various challenges, primarily due to limited diagnostic and control methods. Bacterial culture and acid-fast staining are the standard confirmation methods; however, these procedures are slow and insensitive enough to detect subclinical cases (Jeyanathan et al., 2006). Serological methods and the tuberculin test have limited accuracy, whereas pharmacological treatment in poultry is rarely effective (Roller et al., 2020). Control strategies still rely heavily on biosecurity and environmental management (Ngom et al., 2024). Furthermore, genetic variation in MAA adds complexity, affecting virulence, immune responses, and experimental vaccine effectiveness (González-Pérez et al., 2013). However, advances in genomic technology, molecular surveillance, and diagnostic innovations present new opportunities for ATB research. Bacterial genomic analysis allows for strain identification, understanding of pathogenesis pathways, and development of more specific vaccines or diagnostic biomarkers (Tiwari et al., 2025). Surveillance using PCR and other molecular techniques can improve early detection, map epidemiological distribution, and support risk-based control strategies (Lv et al., 2026). Furthermore, modern immunological approaches and the use of immunomodulators can strengthen the defense response of poultry to infection (Yu et al., 2021). Further research incorporating epidemiology, pathogenesis, genomics, and immunology, and assessing control strategies applicable to commercial farms, is needed (Marzouk and Alajaji, 2025). Interdisciplinary studies are also crucial for evaluating zoonotic risks, economic impacts, and vaccine or biosecurity intervention effectiveness (Belay et al., 2017). A more comprehensive understanding will hopefully lead to the development of faster diagnostic methods, more effective control measures, and sustainable prevention strategies, thereby reducing the disease burden in poultry and minimizing human health risks. ConclusionATB, caused by MAA, is a chronic bird disease with a global distribution and zoonotic potential, particularly in immunocompromised individuals. The disease has significant economic impacts, including reduced poultry productivity, increased mortality, and costs associated with control measures. Second, accurate diagnosis requires a multimodal approach that combines clinical examination, histopathology, bacteriological culture, and molecular techniques, while pharmacological treatments remain largely ineffective. Third, effective control relies on integrated strategies, including strict biosecurity, environmental management, infected bird isolation, and preventive vaccination. Finally, a comprehensive understanding of the epidemiology, diagnostic challenges, and control strategies of ATB is essential for veterinary practice, disease management, and public health safeguarding. AcknowledgmentsThe authors would like to thank the Universitas Airlangga and Badan Riset dan Inovasi Nasional. Conflict of interestThe authors declare no conflict of interest. FundingThis study was supported by the Badan Riset dan Inovasi Nasional, Indonesia, who provided funding for this study by Penerima Program Riset dan Inovasi Untuk Indonesia Maju, year 2023, with grant number: 37/II.7/HK/2023. Author’s contributionsY.P., A.R.K., H.M.R., and W.T. drafted the manuscript. D.A.A.K., I.B.M., B.W.K.W., and F.E. revise and edit the manuscript. B.P.P., S.R., R.Z.A., and I.F.M. prepared and critically checked this manuscript. M.A., F.U., S.S.P., and S.W. edit the references. All authors have read and approved the final version of the manuscript. Data availabilityAll references are open access, so data can be obtained from the internet. ReferencesAbd El-Ghany, W.A. 2022. A review of avian mycobacteriosis: an emerging bacterial disease of public health concern. Int. J. One. Health. 8(2), 70–75. Abukhalid, N., Islam, S., Ndzeidze, R. and Bermudez, L.E. 2021. Mycobacterium avium Subsp. hominissuis interactions with macrophage killing mechanisms. Pathogens 10(11), 1365. Acharya, N., Chattopadhyay, A., Sharma, K. and Sharma, A. 2018. Splenic granulomas: a rare manifestation of Mycobacterium avium complex in an immunocompetent host. BMJ. Case. Rep. 1(1), bcr2018225788. Achkar, J.M. and Casadevall, A. 2013. Antibody-mediated immunity against tuberculosis: implications for vaccine development. Cell. Host. Microbe. 13(3), 250–262. Agdestein, A., Johansen, T.B., Kolbjørnsen, Ø., Jørgensen, A., Djønne, B. and Olsen, I. 2012. A comparative study of Mycobacterium avium subsp. avium and Mycobacterium avium subsp. hominissuis in experimentally infected pigs. BMC. Vet. Res. 8(1), 11. Agdestein, A., Jones, A., Flatberg, A., Johansen, T.B., Heffernan, I.A., Djønne, B., Bosco, A. and Olsen, I. 2014. Intracellular growth of Mycobacterium avium subspecies and global transcriptional responses in human macrophages after infection. BMC. Genomics 15(1), 58. Algammal, A.M., Hashem, H.R., Al-Otaibi, A.S., Alfifi, K.J., El-Dawody, E.M., Mahrous, E., Hetta, H.F., El-Kholy, A.W., Ramadan, H. and El-Tarabili, R.M. 2021. Emerging MDR-Mycobacterium avium subsp. avium in house-reared domestic birds as the first report in Egypt. BMC. Microbiol. 21(1), 237. Angkwanish, T., Vernooij, H.J.C.M., Sirimalaisuwan, A., Charernpan, P., Nielen, M. and Rutten, V.P.M.G. 2021. Prevalence and demographic risk factors of Mycobacterium tuberculosis infections in captive Asian elephants (Elephas maximus) based on serological assays. Front. Vet. Sci. 8(1), 713663. Awuh, J.A. and Flo, T.H. 2017. Molecular basis of mycobacterial survival in macrophages. Cell. Mol. Life Sci. 74(9), 1625–1648. Bao, Y., Yang, T., Zhang, H., Wang, Z., Muhammad, I., Jiang, X. and Ma, H. 2023. Identification and phylogenetic analysis of Mycobacterium avium subsp. avium strain isolated from cow. Transbound. Emerg. Dis. 1(1), 5384079. Baratov, M.O. and Mustafayev, A.R. 2024. Scientific justification of Mycobacterium avium survival in natural environment of Republic of Dagestan. Vet. Sci. Today 13(1), 73–77. Barletta, R.G., Bannantine, J.P., Stabel, J.R., Muthukrishnan, E., Anderson, D.K., Dutta, E., Manthena, V., Hanafy, M. and Zinniel, D.K. 2023. Mycobacterium avium subsp. paratuberculosis candidate vaccine strains are pro-apoptotic in RAW 264.7 murine macrophages. Vaccines 11(6), 1085. Basler, T., Brumshagen, C., Beineke, A., Goethe, R. and Bäumer, W. 2013. Mycobacterium avium subspecies impair dendritic cell maturation. Innate. Immun. 19(5), 451–461. Batista, L.A.F., Silva, K.J.S., da Costa E. Silva., De Moura, Y.F. and Zucchi, F.C.R. 2020. Tuberculosis: a granulomatous disease mediated by epigenetic factors. Tuberculosis (Edinb) 123(1), 101943. Belay, E.D., Kile, J.C., Hall, A.J., Barton-Behravesh, C., Parsons, M.B., Salyer, S. and Walke, H. 2017. Zoonotic disease programs for enhancing global health security. Emerg. Infect. Dis. 23(13), S65–S70. Bielanski, A., Algire, J., Randall, G.C.B. and Surujballi, O. 2006. Risk of transmission of Mycobacterium avium ssp. paratuberculosis by embryo transfer of in vivo and in vitro fertilized bovine embryos. Theriogenology 66(2), 260–266. Blanco Vázquez, C., Alonso-Hearn, M., Juste, R.A., Canive, M., Iglesias, T., Iglesias, N., Amado, J., Vicente, F., Balseiro, A. and Casais, R. 2020. Detection of latent forms of Mycobacterium avium subsp. paratuberculosis infection using host biomarker-based ELISAs greatly improves paratuberculosis diagnostic sensitivity. PLoS One 15(9), e0236336. Borham, M., Oreiby, A., El-Gedawy, A., Hegazy, Y., Khalifa, H.O., Al-Gaabary, M. and Matsumoto, T. 2022. Review on bovine tuberculosis: an emerging disease associated with multidrug-resistant Mycobacterium species. Pathogens 11(7), 715. Boroojeni, A.M., Veiskarami, N., Simula, E.R., Sechi, L.A. and Derakhshandeh, A. 2025. Humoral immune activation against Mycobacterium avium subsp. paratuberculosis through oral immunization with engineered Salmonella. Bacteria 4(3), 47. Butucel, E., Balta, I., Mccleery, D., Morariu, F., Pet, I., Popescu, C.A., Stef, L. and Corcionivoschi, N. 2022. Farm biosecurity measures and interventions with an impact on bacterial biofilms. Agriculture 12(8), 1251. Buur, J. and Saggese, M.D. 2012. Taking a rational approach in the treatment of avian mycobacteriosis. Vet. Clin. North. Am. Exot. Anim. Pract. 15(1), 57–70. Carabalí-Isajar, M.L., Rodríguez-Bejarano, O.H., Amado, T., Patarroyo, M.A., Izquierdo, M.A., Lutz, J.R. and Ocampo, M. 2023. Clinical manifestations and immune response to tuberculosis. World J. Microbiol. Biotechnol. 39(8), 206. Caulfield, A.J. and Wengenack, N.L. 2016. Diagnosis of active tuberculosis disease: from microscopy to molecular techniques. J. Clin. Tuberc. Other. Mycobact. Dis. 4(1), 33–43. Charrier, E.S.A., Dassonville-Klimpt, A., Andréjak, C. and Sonnet, P. 2025. M. avium complex pulmonary infections: therapeutic obstacles and progress in drug development. Pharmaceuticals (Basel) 18(6), 891. Chui, L.W., King, R., Chow, E.Y. and Sim, J. 2004. Immunological response to Mycobacterium avium subsp. paratuberculosis in chickens. Can. J. Vet. Res. 68(4), 302–308. Clark, S., Daly, R., Jordan, E., Lee, J., Mathew, A. and Ebner, P. 2012. Extension education symposium: the future of biosecurity and antimicrobial use in livestock production in the United States and the role of extension. J. Anim. Sci. 90(8), 2861–2872. Coleman, M., Martinez, L., Theron, G., Wood, R. and Marais, B. 2022. Mycobacterium tuberculosis transmission in high-incidence settings-new paradigms and insights. Pathogens 11(11), 1228. Cook, G.M., Berney, M., Gebhard, S., Heinemann, M., Cox, R.A., Danilchanka, O. and Niederweis, M. 2009. Physiology of mycobacteria. Adv. Microbial Physiol. 55(1), 81–182. Corn, J.L., Manning, E.J., Sreevatsan, S. and Fischer, J.R. 2005. Isolation of Mycobacterium avium subsp. paratuberculosis from free-ranging birds and mammals on livestock premises. Appl. Environ. Microbiol. 71(11), 6963–6967. Cromie, R.L., Brown, M.J., Price, D.J. and Stanford, J.L. 1991. Susceptibility of captive wildfowl to avian tuberculosis: the importance of genetic and environmental factors. Tubercle 72(2), 105–109. Daley, C.L. 2017. Mycobacterium avium complex disease. Microbiol. Spectr. 5(2), 663–701; doi: 10.1128/microbiolspec.tnmi7-0045-2017 Dane, H., Koidis, A., Stewart, L.D. and Grant, I.R. 2022. Optimization of the composition of a solid culture medium for Mycobacterium avium subsp. paratuberculosis using factorial design and response surface methodology. J. Appl. Microbiol. 132(6), 4252–4265. Danelishvili, L., Armstrong, E., Miyasako, E., Jeffrey, B. and Bermudez, L.E. 2020. Exposure of Mycobacterium avium subsp. homonissuis to metal concentrations of the phagosome environment enhances the selection of persistent subpopulation to antibiotic treatment. Antibiotics (Basel) 9(12), 927. Dao, T.O., Park, H.-E., Lee, J.H., Kim, K.-M., Trinh, M.P., Kang, H.-L., Yoo, H.S. and Shin, M.-K. 2025. Advances and challenges in mycobacterial genetic engineering: techniques for knockout, knockdown and overexpression. J. Microbiol. Biotechnol. 35(1), e2507051. Darya, B.K., Nader, M. and Kaveh, P.A. 2025. Comparative study of clinical examinations, necropsy findings, and X-ray signs in naturally infected pigeons to avian tuberculosis. Arch. Razi. Inst. 80(2), 509–516. De Lisle, G.W., Wards, B.J., Buddle, B.M. and Collins, D.M. 2005. The efficacy of live tuberculosis vaccines after presensitization with Mycobacterium avium. Tuberculosis (Edinb) 85(1–2), 73–79. Debelu, T., Abunna, F. and Kassa, G.M. 2021. A preliminary study on public health implications of avian tuberculosis in selected districts of the Oromia Region, Ethiopia. Vet. Med. Int. 2021(1), 6331599. Dhama, K., Mahendran, M., Tiwari, R., Singh, S.D., Kumar, D., Singh, S. and Sawant, P.M. 2011. Tuberculosis in birds: insights into the Mycobacterium avium infections. Vet. Med. Int. 2011(1), 712369. Dvorska, L., Matlova, L., Ayele, W.Y., Fischer, O.A., Amemori, T., Weston, R.T., Alvarez, J., Beran, V., Moravkova, M. and Pavlik, I. 2007. Avian tuberculosis in naturally infected captive water birds of the Ardeideae and Threskiornithidae families studied by serotyping, IS901 RFLP typing, and virulence for poultry. Vet. Microbiol. 119(2–4), 366–374. Elmeligy, A., Ghania, A. and Fotouh, A. 2024. Pathological and immunohistochemical studies of lymphoid leukosis in pigeons in Egypt. Open Vet. J. 14(8), 1952–1959. Falkinham, J.O. 2003. Factors influencing the chlorine susceptibility of Mycobacterium avium, Mycobacterium intracellulare, and Mycobacterium scrofulaceum. Appl. Environ. Microbiol. 69(9), 5685–5689. Fernandez-Pittol, M., Batista-Arnau, S., Román, A., San Nicolás, L., Oliver, L., González-Moreno, O., Martínez, J.A., Amaro-Rodríguez, R., Soler, N., Gené, A., González-Cuevas, A., Tudó, G. and Gonzalez-Martin, J. 2022. Differences in drug-susceptibility patterns between Mycobacterium avium, Mycobacterium intracellulare, and Mycobacterium chimaera clinical isolates: prospective 8.5-year analysis by three laboratories. Antibiotics (Basel) 12(1), 64. Ferreira, E.M., Cunha, M.V., Duarte, E.L., Gonçalves, R., Pinto, T., Mira, A. and Santos, S.M. 2024. Host-, environment-, or human-related effects drive interspecies interactions in an animal tuberculosis multi-host community depending on the host and season. Transbound. Emerg. Dis. 2024(1), 9779569. Flórido, M. and Appelberg, R. 2004. Granuloma necrosis during Mycobacterium avium infection does not require tumor necrosis factor. Infect. Immun. 72(10), 6139–6141. Forrellad, M.A., Klepp, L.I., Gioffré, A., Sabio Y García, J., Morbidoni, H.R., Santangelo, M.D.L.P., Cataldi, A.A. and Bigi, F. 2013. Virulence factors of the Mycobacterium tuberculosis complex. Virulence 4(1), 3–66. Garvey, M. 2020. Mycobacterium avium paratuberculosis: a disease burden on the dairy industry. Animals (Basel) 10(10), 1773. Gill, C.O., Saucier, L. and Meadus, W.J. 2011. Mycobacterium avium subsp. paratuberculosis in dairy products, meat, and drinking water. J. Food Protect. 74(3), 480–499. Gomes, M.S., Paul, S., Moreira, A.L., Appelberg, R., Rabinovitch, M. and Kaplan, G. 1999. Survival of Mycobacterium avium and Mycobacterium tuberculosis in acidified vacuoles of murine macrophages. Infect. Immun. 67(7), 3199–3206. González-Pérez, M., Mariño-Ramírez, L., Parra-López, C.A., Murcia, M.I., Marquina, B., Mata-Espinoza, D., Rodriguez-Míguez, Y., Baay-Guzman, G.J., Huerta-Yepez, S. and Hernandez-Pando, R. 2013. Virulence and immune response induced by Mycobacterium avium complex strains in a model of progressive pulmonary tuberculosis and subcutaneous infection in BALB/c mice. Infect. Immun. 81(11), 4001–4012. Grace, D., Knight-Jones, T.J.D., Melaku, A., Alders, R. and Jemberu, W.T. 2024. The public health importance and management of infectious poultry diseases in smallholder systems in Africa. Foods 13(3), 411. Grewal, S.K., Rajeev, S., Sreevatsan, S. and Michel, F.C. 2006. Persistence of Mycobacterium avium subsp. paratuberculosis and other zoonotic pathogens during simulated composting, manure packing, and liquid storage of dairy manure. Appl. Environ. Microbiol. 72(1), 565–574. Guarner, J. and Brandt, M.E. 2011. Histopathologic diagnosis of fungal infections in the 21st century. Clin. Microbiol. Rev. 24(2), 247–280. Gupta, S.K., Wilson, T., Maclean, P.H., Rehm, B.H.A., Heiser, A., Buddle, B.M. and Wedlock, D.N. 2023. Mycobacterium avium subsp. paratuberculosis antigens induce cellular immune responses in cattle without causing reactivity to tuberculin in the tuberculosis skin test. Front. Immunol. 13(1), 1087015. Haque, M.N., Rima, U.K., Hossain, M.Z., Islam, M.S., Chowdhury, S.M.Z.H., Hossain, M.M. and Khan, M.A.H.N.A. 2016. Standardize polymerase chain reaction (PCR) technique for the detection of pathogenic serovars of Mycobacterium a. avium infection in layer chicken. Asian J. Poult. Sci. 10(4), 175–183. Harris, N.B. and Barletta, R.G. 2001. Mycobacterium avium subsp. paratuberculosis in veterinary medicine. Clin. Microbiol. Rev. 14(3), 489–512. Hasan, M.K., Rahman, M.M., Kisku, J.J., Rahman, M.B., Rahman, M.M. and Rahman, M.M. 2016. Prevalence of bovine and avian tuberculosis in calves of central cattle breeding and dairy farm, Savar, Dhaka. J. Innov. Dev. Strategy. 10(2), 5–12. Hernández, A.J.C. 2014. In Encyclopedia of Agriculture and Food Systems. Ed., Neal K. Van Alfen. Amsterdam, The Netherlands: Elsevier, pp: 504–520. Hernández-Agudelo, J.M., Verdugo, C., Barkema, H.W., Steuer, P., Tejeda, C., Ulloa, F., Soto, J.P. and Salgado, M.A. 2025. Early-life shedding and environmental presence of Mycobacterium avium ssp. paratuberculosis in chilean dairy calves. J. Dairy. Sci. 108(11), 12605–12617. Howard, C.J., Kwong, L.S., Villarreal-Ramos, B., Sopp, P. and Hope, J.C. 2002. Exposure to Mycobacterium avium primes the immune system of calves for vaccination with Mycobacterium bovis BCG. Clin. Exp. Immunol. 130(2), 190–195. Ignatov, D., Kondratieva, E., Azhikina, T. and Apt, A. 2012. Mycobacterium avium-triggered diseases: pathogenomics. Cell. Microbiol. 14(6), 808–818. Jeyanathan, M., Alexander, D.C., Turenne, C.Y., Girard, C. and Behr, M.A. 2006. Evaluation of in situ methods used to detect Mycobacterium avium subsp. paratuberculosis in samples from patients with Crohn’s disease. J. Clin. Microbiol. 44(8), 2942–2950. Joseph, J.J., Leestemaker-Palmer, A., Kazemi, S., Danelishvili, L. and Bermudez, L.E. 2023. Mycobacterium avium infection of multinucleated giant cells reveals association of bacterial survival to autophagy and cholesterol utilization. Cell. Microbiol. 1(1), 5064371. Kaboudi, K., Amara, A. and Bouzouaia, M. 2017. Avian tuberculosis in a backyard poultry flock in Tunisia: case report. Int. J. Vet. Sci. Anim. Husb. 2(6), 34–37. Kaevska, M., Slana, I., Kralik, P. and Pavlik, I. 2010. Examination of Mycobacterium avium subsp. avium distribution in naturally infected hens by culture and triplex quantitative real time PCR. Vet. Med. 55(7), 325–330. Kaiser, P. 2010. Advances in avian immunology--prospects for disease control: a review. Avian. Pathol. 39(5), 309–324. Kang, X., Yang, Y., Meng, C., Wang, X., Liu, B., Geng, S., Jiao, X. and Pan, Z. 2022. Safety and protective efficacy of Salmonella pullorum spiC and rfaH deletion rough mutant as a live attenuated DIVA vaccine candidate. Poult. Sci. 101(3), 655. Kannan, N., Lai, Y.P., Haug, M., Lilleness, M.K., Bakke, S.S., Marstad, A., Hov, H., Naustdal, T., Afset, J.E., Ioerger, T.R., Flo, T.H. and Steigedal, M. 2019. Genetic variation/evolution and differential host responses resulting from in-patient adaptation of Mycobacterium avium. Infect. Immun. 87(4), 23. Khairullah, A., Moses, I., Kusala, M., Tyasningsih, W., Ayuti, S., Rantam, F., Fauziah, I., Silaen, O., Puspitasari, Y., Aryaloka, S., Raharjo, H., Hasib, A., Yanestria, S. and Nurhidayah, N. 2024. Unveiling insights into bovine tuberculosis: a comprehensive review. Open Vet. J. 14(6), 1330–1344. Kim, S., Park, H.E., Park, W.B., Kim, S.Y., Park, H.T. and Yoo, H.S. 2021. Mycobacterium avium modulates the protective immune response in canine peripheral blood mononuclear cells. Front. Cell. Infect. Microbiol. 10(1), 60971. Kindu, A. and Getaneh, G. 2016. Prevalence of avian tuberculosis in domestic chickens in selected sites of Ethiopia. J. Vet. Sci. Technol. 7(6), 377. Knific, T., Ocepek, M., Kirbiš, A., Krt, B., Prezelj, J. and Gethmann, J.M. 2022. Quantitative risk assessment of exposure to Mycobacterium avium subsp. paratuberculosis (MAP) via different types of milk for the Slovenian consumer. Foods 11(10), 1472. Kobuszewska, A. and Wysok, B. 2024. Pathogenic bacteria in free-living birds, and its public health significance. Animals 14(6), 968. Koets, A.P., Eda, S. and Sreevatsan, S. 2015. The within host dynamics of Mycobacterium avium ssp. paratuberculosis infection in cattle: where time and place matter. Vet. Res. 46(1), 61. Komatsu, T., Inaba, N., Kondo, K., Nagata, R., Kawaji, S. and Shibahara, T. 2017. Systemic mycobacteriosis caused by ‘Mycobacterium avium subspecies hominissuis’ in a 14-month-old Japanese black beef steer. J. Vet. Med. Sci. 79(8), 1384–1388. Koo, H.C., Park, Y.H., Hamilton, M.J., Barrington, G.M., Davies, C.J., Kim, J.B., Dahl, J.L., Waters, W.R. and Davis, W.C. 2004. Analysis of the immune response to Mycobacterium avium subsp. paratuberculosis in experimentally infected calves. Infect. Immun. 72(12), 6870–6883. Kriz, P., Kaevska, M., Bartejsova, I. and Pavlik, I. 2013. Mycobacterium avium subsp. avium found in raptors exposed to infected domestic fowl. Avian. Dis. 57(3), 688–692. Kubota, Y., Takasawa, A., Ono, Y., Aoyama, T., Takasawa, K., Tada, A., Magara, K., Murakami, T., Daimon, F., Yamamoto, S., Sato, S., Hiratsuka, Y., Kyuno, D. and Osanai, M. 2023. Invasive pulmonary aspergillosis with candidiasis: usefulness of molecular and ultrastructural morphological analysis on FFPE tissue for invasive fungal infections. Med. Mol. Morphol. 56(2), 144–151. Kwaghe, A.V., Bello, M., Balogun, M.S., Audu, B.J. and Kabir, J. 2024. The use of participatory disease surveillance, tuberculin test and polymerase chain reaction to determine the presence and prevalence of avian tuberculosis in layers within Gwagwalada and Kuje Area Councils, Abuja, Nigeria: a cross-sectional study. Arch. Microbiol. Immunol. 8(1), 501–512. Kwaghe, A.V., Ndahi, M.D., Usman, J.G., Vakuru, C.T., Iwar, V.N., Ameh, J.A. and Ambali, A.G. 2015. A review on avian tuberculosis (Avian mycobacteriosis). CAB Rev. 10, 1–17. Lâm, S., Dang-Xuan, S., Unger, F., Meeyam, T., Pham-Duc, P., Wacharapluesadee, S. and Nguyen-Viet, H. 2025. Operationalizing regional One Health initiatives in Southeast Asia: ways forward. One Health 20(1), 101034. Lari Baghal, M., Mayahi, M., Mosavari, N., Boroomand, Z. and Islampanah, M. 2024. Comparison of isolation and histopathology methods in diagnosis of pigeon tuberculosis. J. Poult. Sci. Avian. Dis. 2(1), 27–35. Lechevallier, M.W., Prosser, T. and Stevens, M. 2024. Opportunistic pathogens in drinking water distribution systems—a review. Microorganisms 12(5), 916. Ledwoń, A., Dolka, I., Dolka, B., Cegiełkowska, M., Czopowicz, M. and Szeleszczuk, P. 2015. Multidrug therapy of Mycobacterium avium subsp. avium infection in experimentally inoculated budgerigars (Melopsittacus undulatus). Avian Pathol. 44(6), 470–474. Lekko, Y., Che-Amat, A., Ooi, P., Omar, S., Ramanoon, S., Mazlan, M., Jesse, F., Jasni, S. and Ariff Abdul-razak, M. 2021. Mycobacterium tuberculosis and avium complex investigation among Malaysian free-ranging wild boar and wild Macaques at wildlife-livestock-human interface. Animals 11(11), 3252. Levison, M.E. 2015. Diseases transmitted by birds. Microbiol. Spectr. 3(4), IOL5-0004-2015. Li, H., Ge, M. and Zhang, M. 2022. Spatio-temporal distribution of tuberculosis and the effects of environmental factors in China. BMC. Infect. Dis. 22(1), 565. Li, Y.J., Danelishvili, L., Wagner, D., Petrofsky, M. and Bermudez, L.E. 2010. Identification of virulence determinants of Mycobacterium avium that impact on the ability to resist host killing mechanisms. J. Med. Microbiol. 59(Pt 1), 8–16. Libera, K., Konieczny, K., Grabska, J., Szopka, W., Augustyniak, A. and Pomorska-Mól, M. 2022. Selected livestock-associated zoonoses as a growing challenge for public health. Infect. Dis. Rep. 14(1), 63–81. Liu, H., Pan, S., Wang, C., Yang, W., Wei, X., He, Y., Xu, T., Shi, K. and Si, H. 2025. Review of respiratory syndromes in poultry: pathogens, prevention, and control measures. Vet. Res. 56(1), 101. Lombard, J.E. and Garry, F.B. 2025. Biosecurity practices for Mycobacterium avium subspecies paratuberculosis infection, salmonellosis, and bovine leukemia virus on cattle operations. Vet. Clin. North Am. Food Anim. Pract. 41(1), 93–101. Lv, Y.R., Liu, Y.Y., Zhang, R., Yang, B., Xue, S.Y., Ding, Y.L., Jia, J.T., Bayaer, H., Bagen, A., Chen, R.B., Tunala, S., Zhao, L. and Liu, Y.H. 2026. Rapid and simple detection of Mycobacterium avium subsp. paratuberculosis using a lateral flow assay based on CRISPR-Cas12a combined with recombinase polymerase amplification or nested PCR. Pathogens 15(1), 24. Lyu, J., Narum, D.E., Baldwin, S.L., Larsen, S.E., Bai, X., Griffith, D.E., Dartois, V., Naidoo, T., Steyn, A.J.C., Coler, R.N. and Chan, E.D. 2024. Understanding the development of tuberculous granulomas: insights into host protection and pathogenesis, a review in humans and animals. Front. Immunol. 15(1), 1427559. Mai, T.N., Nguyen, T.T., Dang-Xuan, S., Nguyen-Viet, H., Unger, F. and Lee, H.S. 2024. Transboundary viral diseases of pigs, poultry and ruminants in Southeast Asia: a systematic review. Vet. Q. 44(1), 13–25. Maina, J.N. 2023. A critical assessment of the cellular defences of the avian respiratory system: are birds in general and poultry in particular relatively more susceptible to pulmonary infections/afflictions?. Biol. Rev. 98(6), 2152–2187. Mamo, G. 2020. Abattoir-based prevalence of avian tuberculosis in chicken slaughtered at poultry abattoir in Bishoftu, Central Ethiopia. Ethiop. Vet. J. 24(1), 1–14. Mariappan, A.K., Mathesh, K., Muthu, S., Bhatt, M., Sharma, M., Saikumar, G. and Dhama, K. 2023. Pathological and molecular identification of Mycobacterium avium infection in a loft of domestic pigeons (Columba livia var. domestica) from India. Braz. J. Microbiol. 54(3), 2521–2526. Martin, G. and Schimmel, D. 2000. Die Mycobacterium avium-Infektion des Geflügels--(k)eine Gefahr für die menschliche Gesundheit? Mycobacterium avium infections in poultry--a risk for human health or not?. Dtsch. Tierarztl. Wochenschr. 107(2), 53–58. Marzouk, E. and Alajaji, A.I. 2025. Preventive immunology for livestock and zoonotic infectious diseases in the One Health Era: from mechanistic insights to innovative interventions. Vet. Sci. 12(10), 1014. Matos, S., Portugal, I. and Perdigão, J. 2025. The Mycobacterium avium complex: genomics, disease, and beyond. Microorganisms 13(10), 2329. Mayahi, M., Esmaeilzadeh, S., Mosavari, N. and Asadollahi, K.P. 2013a. Histopathological study of avian tuberculosis in naturally infected domestic pigeons with Mycobacterium avium subsp. avium. Iran. J. Vet. Sci. Technol. 5(1), 45–56. Mayahi, M., Mosavari, N., Esmaeilzadeh, S. and Asadollahi, K.P. 2013b. Avian tuberculosis in naturally infected lofts of domestic pigeons, isolation, molecular identification and study of necropsy findings. Intern. J. Appl. Res. Vet. Med. 11(3), 194–201. Mehmedi, B., Iatrou, A.M., Yildiz, R., Lamont, K., Da Costa, M.R., De Nardi, M., Allepuz, A., Niine, T., Niemi, J.K. and Saegerman, C. 2025. Economic perspectives on farm biosecurity: stakeholder challenges and livestock species considerations. Agriculture 15(21), 2288. Mendoza-Trujillo, I., Diez-Echave, P., Tontini, C. and Bulfone-Paus, S. 2025. The crosstalk between Mycobacterium abscessus and immune cells: exploring novel interaction modalities. Cells 14(22), 1829. Miggiano, R., Rizzi, M. and Ferraris, D.M. 2020. Mycobacterium tuberculosis pathogenesis, infection prevention and treatment. Pathogens 9(5), 385. Moravkova, M., Lamka, J., Kriz, P. and Pavlik, I. 2011. The presence of Mycobacterium avium subsp. avium in common pheasants (Phasianus colchicus) living in captivity and in other birds, vertebrates, non-vertebrates and the environment. Vet. Med. 56(7), 333–343. Motiwala, A.S., Strother, M., Amonsin, A., Byrum, B., Naser, S.A., Stabel, J.R., Shulaw, W.P., Bannantine, J.P., Kapur, V. and Sreevatsan, S. 2003. Molecular epidemiology of Mycobacterium avium subsp. paratuberculosis: evidence for limited strain diversity, strain sharing, and identification of unique targets for diagnosis. J. Clin. Microbiol. 41(5), 2015–2026. Ncube, P., Bagheri, B., Goosen, W.J., Miller, M.A. and Sampson, S.L. 2022. Evidence, challenges, and knowledge gaps regarding latent tuberculosis in animals. Microorganisms 10(9), 1845. Nesic, V., Marinkovic, D., Matovic, K., Radakovic, M., Davitkov, D., Vaskovic, N. and Davitkov, D. 2022. Avian tuberculosis in a free-living Eurasian griffon vulture. J. Vet. Diagn. Invest. 34(4), 723–726. Ngom, R.V., Ayissi, G.J., Akoussa, A.M.M., Laconi, A., Jajere, S.M., Zangue, H.A. and Piccirillo, A. 2024. A systematic review and meta-analysis of the efficacy of biosecurity in disease prevention and control in livestock farms in Africa. Transbound. Emerg. Dis. 2024(1), 8683715. Nhung, N.T., Chansiripornchai, N. and Carrique-Mas, J.J. 2017. Antimicrobial resistance in bacterial poultry pathogens: a review. Front. Vet. Sci. 4(1), 126. Nielsen, S.S., Kolmos, B. and Christoffersen, A.B. 2004. Comparison of contamination and growth of Mycobacterium avium subsp. paratuberculosis on two different media. J. Appl. Microbiol. 96(1), 149–153. Olowu, B.I., Zakariya, M.E., Azeez, A.O., Al-Awal, A.A., Adebayo, K.S., Idris, N.O., Muhammad, H.I., Ukauwa, B.C. and Olojede, A.A.A. 2026. Propagation of emerging and re-emerging infectious disease pathogens in Africa: the role of migratory birds. Bacteria 5(1), 2. Orujyan, D., Narinyan, W., Rangarajan, S., Rangchaikul, P., Prasad, C., Saviola, B. and Venketaraman, V. 2022. Protective efficacy of BCG vaccine against Mycobacterium leprae and non-tuberculous mycobacterial infections. Vaccines 10(3), 390. Park, H.E., Kim, K.M., Trinh, M.P., Yoo, J.W., Shin, S.J. and Shin, M.K. 2024. Bigger problems from smaller colonies: emergence of antibiotic-tolerant small colony variants of Mycobacterium avium complex in MAC-pulmonary disease patients. Ann. Clin. Microbiol. Antimicrob. 23(1), 25. Park, H.E., Lee, W., Choi, S., Jung, M., Shin, M.K. and Shin, S.J. 2022. Modulating macrophage function to reinforce host innate resistance against Mycobacterium avium complex infection. Front. Immunol. 13(1), 931876. Park, H.T. and Yoo, H.S. 2016. Development of vaccines to Mycobacterium avium subsp. paratuberculosis infection. Clin. Exp. Vaccine. Res. 5(2), 108–116. Parvandar-Asadollahi, K., Mosavari, N. and Mayahi, M. 2015. Genotyping of Mycobacterium avium subsp. avium isolates from naturally infected lofts of domestic pigeons in Ahvaz by IS901 RFLP. Iran. J. Microbiol. 7(5), 260–264. Pereira, A.C., Reis, A.C., Ramos, B. and Cunha, M.V. 2020. Animal tuberculosis: impact of disease heterogeneity in transmission, diagnosis and control. Transbound. Emerg. Dis. 67(5), 1828–1846. Rabinowitz, P.M., Kock, R., Kachani, M., Kunkel, R., Thomas, J., Gilbert, J., Wallace, R., Blackmore, C., Wong, D., Karesh, W., Natterson, B., Dugas, R. and Rubin, C. 2013. Toward proof of concept of a one health approach to disease prediction and control. Emerg. Infect. Dis. 19(12), e130265. Rais, M., Abdelaal, H., Reese, V.A., Ferede, D., Larsen, S.E., Pecor, T., Erasmus, J.H., Archer, J., Khandhar, A.P., Cooper, S.K., Podell, B.K., Reed, S.G., Coler, R.N. and Baldwin, S.L. 2023. Immunogenicity and protection against Mycobacterium avium with a heterologous RNA prime and protein boost vaccine regimen. Tuberculosis (Edinb) 138(1), 102302. Rathnaiah, G., Zinniel, D.K., Bannantine, J.P., Stabel, J.R., Gröhn, Y.T., Collins, M.T. and Barletta, R.G. 2017. Pathogenesis, molecular genetics, and genomics of Mycobacterium avium subsp. paratuberculosis, the etiologic agent of Johne’s disease. Front. Vet. Sci. 4(1), 187. Reil, I., Duvnjak, S., Špičić, S., Kompes, G., Bagarić, A., Đuras, M., Gudan Kurilj, A., Lukač, M., Jelić, M. and Zdelar-Tuk, M. 2024. Isolation of multidrug-resistant Mycobacterium avium Subsp. avium from a Wild Eurasian Otter (Lutra lutra). Antibiotics 13(7), 591. Reza, M., Lijon, M., Khatun, M. and Islam, M. 2015. Prevalence and antibiogram profile of Mycobacterium spp. in poultry and its environments. J. Adv. Vet. Anim. Res. 2(4), 458–463. Riber, A.B. and Wurtz, K.E. 2024. Impact of growth rate on the welfare of broilers. Animals 14(22), 3330. Roberts, J.R., Souillard, R. and Bertin, J. 2011. Avian diseases which affect egg production and quality. Improving Saf. Qual. Eggs Egg Prod. 1(1), 376–393. Roller, M., Hansen, S., Knauf-Witzens, T., Oelemann, W.M.R., Czerny, C.P., Abd El Wahed, A. and Goethe, R. 2020. Mycobacterium avium subspecies paratuberculosis infection in zoo animals: a review of susceptibility and disease process. Front. Vet. Sci. 7(1), 572724. Roque, S., De Sá-calçada, D., Cerqueira-Rodrigues, B., Monteiro, S., Guerreiro, S.G., Palha, J.A. and Correia-Neves, M. 2023. Chronic Mycobacterium avium infection differentially affects the cytokine expression profile of three mouse strains, but has no effect on behavior. Sci. Rep. 13(1), 6199. Roy, S., Ghatak, D., Das, P. and BoseDasgupta, S. 2020. ESX secretion system: the gatekeepers of mycobacterial survivability and pathogenesis. Eur. J. Microbiol. Immunol. (Bp). 10(4), 202–209. Saggese, M.D., Tizard, I., Gray, P. and Phalen, D.N. 2014. Evaluation of multidrug therapy with azithromycin, rifampin, and ethambutol for the treatment of Mycobacterium avium subsp avium in Ring-neck Doves (Streptopelia risoria): an uncontrolled clinical study. J. Avian. Med. Surg. 28(4), 280–289. Salamatian, I., Ghaniei, A., Mosavari, N., Nourani, H., Keshavarz, R. and Eslampanah, M. 2020. Outbreak of avian mycobacteriosis in a commercial turkey breeder flock. Avian Pathol. 49(3), 296–304. Sattar, A., Zakaria, Z., Abu, J., Aziz, S.A. and Rojas-Ponce, G. 2021. Isolation of Mycobacterium avium and other nontuberculous mycobacteria in chickens and captive birds in peninsular Malaysia. BMC Vet. Res. 17(1), 13. Sawaswong, V., Wongjarit, K., Petsong, S., Yuliani, Y., Somsukpiroh, U., Faksri, K., Forde, T., Payungporn, S. and Rotcheewaphan, S. 2025. Diversity and antimicrobial resistance profiles of Mycobacterium avium complex clinical isolates in Thailand based on whole genome comparative analysis. Sci. Rep. 15(1), 772. Schinköthe, J., Köhler, H. and Liebler-Tenorio, E.M. 2016. Characterization of tuberculous granulomas in different stages of progression and associated tertiary lymphoid tissue in goats experimentally infected with Mycobacterium avium subsp. hominissuis. Comparative. Immunol. Microbiol. Infect. Dis. 47(1), 41–51. Schmidt, V., Köhler, H., Heenemann, K. and Möbius, P. 2022. Mycobacteriosis in various pet and wild birds from Germany: pathological Findings, coinfections, and characterization of causative mycobacteria. Microbiol. Spectr. 10(4), 45222. Severova, L., Giller, D., Enilenis, I., Gadzhieva, P., Shcherbakova, G., Kesaev, O., Koroev, V., Frolova, O., Popova, A., Ilyukhin, A., Basangova, V., Belova, E., Gazi, E.P., Taushkanova, I. and Martel, I. 2025. Detection, isolation, and identification of mycobacteria that cause nontuberculous mycobacterial disease and tuberculosis. Pathogens 14(12), 1302. Sevilla, I.A., Molina, E., Elguezabal, N., Pérez, V., Garrido, J.M. and Juste, R.A. 2015. Detection of mycobacteria, Mycobacterium avium subspecies, and Mycobacterium tuberculosis complex by a novel tetraplex real-time PCR assay. J. Clin. Microbiol. 53(3), 930–940. Shin, M.K. and Shin, S.J. 2021. Genetic involvement of Mycobacterium avium complex in the regulation and manipulation of innate immune functions of host cells. Int. J. Mol. Sci. 22(6), 3011. Shitaye, J.E., Matlova, L., Horvathova, A., Moravkova, M., Dvorska-Bartosova, L., Treml, F., Lamka, J. and Pavlik, I. 2008. Mycobacterium avium subsp. avium distribution studied in a naturally infected hen flock and in the environment by culture, serotyping and IS901 RFLP methods. Vet. Microbiol. 127(1–2), 155–164. Shivaprasad, H.L. and Palmieri, C. 2012. Pathology of mycobacteriosis in birds. Vet. Clin. North Am. Exot. Anim. Pract. 15(1), 41–55. Siddiqui, M.J., Karmacharya, A., Wan, X., Zhu, Y., Wan, C. and Luo, S. 2025. Clinical characteristics leading to misdiagnosis of abdominal tuberculosis in children: a systematic review and meta-analysis. Front. Pediatr. 13(1), 1616608. del Silva A, Leon, C.I., Guerrero, M.I., Neira, R., Arias, L. and Rodriguez, G. 2009. Avian tuberculosis of zoonotic importance at a zoo on the Bogotá Andean plateau (Sabana), Colombia. Can. Vet. J. 50(8), 841–845. Silva, S., Arinaminpathy, N., Atun, R., Goosby, E. and Reid, M. 2021. Economic impact of tuberculosis mortality in 120 countries and the cost of not achieving the sustainable development goals tuberculosis targets: a full-income analysis. Lancet Glob. Health 9(10), e1372–e1379. Singh, A. and Kashyap, V.K. 2012. Specific and rapid detection of Mycobacterium tuberculosis complex in clinical samples by polymerase chain reaction. Interdiscip. Perspect. Infect. Dis. 1(1), 654694. Slany, M., Ulmann, V. and Slana, I. 2016. Avian mycobacteriosis: still existing threat to humans. Biomed. Res. Int. 2016(1), 4387461. Soler, D., Brieva, C. and Ribón, W. 2009. Mycobacteriosis in wild birds: the potential risk of disseminating a little-known infectious disease. Rev. Salud Publica. (Bogota). 11(1), 134–144. Song, Q., Liu, J. and Wang, C. 2026. Tuberculosis: clinical laboratory diagnostic techniques and future perspectives. Vaccines 14(1), 38. Srivastava, V., Dahiya, A., Singh, S.V. and Kulshreshtha, S. 2017. Diagnostic approaches to avian tuberculosis. World’s. Poult. Sci. J. 73(4), 857–871. Ssekitoleko, J., Ojok, L., Abd El Wahed, A., Erume, J., Amanzada, A., Eltayeb, E., Eltom, K.H. and Okuni, J.B. 2021. Mycobacterium avium subsp. paratuberculosis virulence: a review. Microorganisms 9(12), 2623. Stepień-Pyśniak, D., Puk, K., Guz, L., Wawrzyniak, A., Marek, A. and Kosikowska, U. 2016. Avian mycobacteriosis caused by Mycobacterium avium subspecies avium in four ornamental birds and in vitro drug sensitivity testing of isolates. Berl. Münch. Tierärztl. Wochenschr. 129(1–2), 65–71. Tell, L.A., Foley, J., Needham, M.L. and Walker, R.L. 2003. Diagnosis of avian mycobacteriosis: comparison of culture, acid-fast stains, and polymerase chain reaction for the identification of Mycobacterium avium in experimentally inoculated Japanese quail (Coturnix coturnix japonica). Avian Dis. 47(2), 444–452. Tell, L.A., Woods, L. and Cromie, R.L. 2001. Mycobacteriosis in birds. Rev. Sci. Tech. 20(1), 180–203. Thom, M., Howard, C., Villarreal-Ramos, B., Mead, E., Vordermeier, M. and Hope, J. 2008. Consequence of prior exposure to environmental mycobacteria on BCG vaccination and diagnosis of tuberculosis infection. Tuberculosis (Edinb) 88(4), 324–334. Tiwari, S., Dhakal, T., Kim, B.J., Jang, G.S. and Oh, Y. 2025. Genomics in epidemiology and disease surveillance: an exploratory analysis. Life (Basel) 15(12), 1848. To, K., Cao, R., Yegiazaryan, A., Owens, J. and Venketaraman, V. 2020. general overview of nontuberculous mycobacteria opportunistic pathogens: Mycobacterium avium and Mycobacterium abscessus. J. Clin. Med. 9(8), 2541. Tsiouris, V., Kiskinis, K., Mantzios, T., Dovas, C.I., Mavromati, N., Filiousis, G., Brellou, G., Vlemmas, I. and Georgopoulou, I. 2021. Avian mycobacteriosis and molecular identification of Mycobacterium avium Subsp. avium in Racing Pigeons (Columba livia domestica) in Greece. Animals (Basel). 11(2), 291. Turenne, C.Y., Wallace, R. and Behr, M.A. 2007. Mycobacterium avium in the postgenomic era. Clin. Microbiol. Rev. 20(2), 205–229. Verdugo, C., Jones, G., Johnson, W., Wilson, P., Stringer, L. and Heuer, C. 2014. Estimation of flock/herd-level true Mycobacterium avium subspecies paratuberculosis prevalence on sheep, beef cattle and deer farms in New Zealand using a novel Bayesian model. Prev. Vet. Med. 117(3–4), 447–455. Villaamil, F.J., Yus, E., Benavides, B., Allepuz, A., Moya, S.J., Casal, J., Ortega, C. and Diéguez, F.J. 2021. Factors associated with the introduction of Mycobacterium avium spp. paratuberculosis (MAP) into dairy herds in Galicia (North-West Spain): the perception of experts. Animals (Basel) 11(1), 166. Wang, H., Liu, D. and Zhou, X. 2023. Effect of mycolic acids on host immunity and lipid metabolism. Int. J. Mol. Sci. 25(1), 396. Wang, J. and Lee, S. 2025. Targeting autophagy as a strategy for developing new host-directed therapeutics against nontuberculous mycobacteria. Pathogens 14(5), 472. Whittington, R.J., Marshall, D.J., Nicholls, P.J., Marsh, I.B. and Reddacliff, L.A. 2004. Survival and dormancy of Mycobacterium avium subsp. paratuberculosis in the environment. Appl. Environ. Microbiol. 70(5), 2989–3004. Witte, C.L., Hungerford, L.L., Papendick, R., Stalis, I.H. and Rideout, B.A. 2008. Investigation of characteristics and factors associated with avian mycobacteriosis in zoo birds. J. Vet. Diagn. Investig. 20(2), 186–196. Woodbury, M.R., Chirino-Trejo, M. and Mihajlovic, B. 2008. Diagnostic detection methods for Mycobacterium avium subsp. paratuberculosis in white-tailed deer. Can. Vet. J. 49(7), 683–688. Yavuz, O., Özdemir, O., Sayin, Z., Hatipoğlu, F. and Hadimli, H.H. 2021. Diagnosis of avian tuberculosis in laying hens by pathological, microbiological and polymerase chain reaction (PCR): case report. J. Adv. Vet. Bio. Sci. Tech. 6(3), 312–317. Yu, K., Choi, I. and Yun, C.H. 2021. Immunosecurity: immunomodulants enhance immune responses in chickens. Anim. Biosci. 34(3), 321–337. Zhang, C., Wang, L., Zhang, C., Zhang, N., Sun, H., Chu, D., Qin, S., Ma, Z., Gulyaeva, M., Shestopalov, A., Liu, W., Gao, G.F. and Bi, Y. 2025. Avian tuberculosis identified as the potential disease in an outbreak in wild migratory birds in China. mLife 4(1), 101–103. Zhu, D.K., Song, X.H., Wang, J.B., Zhou, W.S., Ou, X.M., Chen, H.X., Liu, M.F., Wang, M.S., Jia, R.Y., Chen, S., Sun, K.F., Yang, Q., Wu, Y., Chen, X.Y. and Cheng, A.C. 2016. Outbreak of avian tuberculosis in commercial domestic Pekin ducks (Anas platyrhynchos domestica). Avian Dis. 60(3), 677–680. | ||
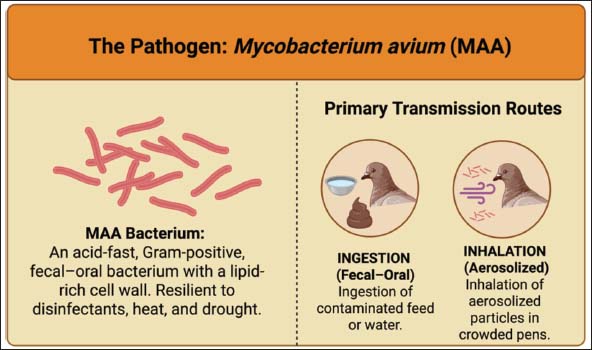
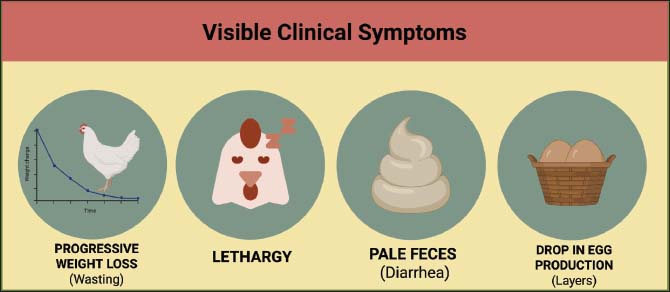
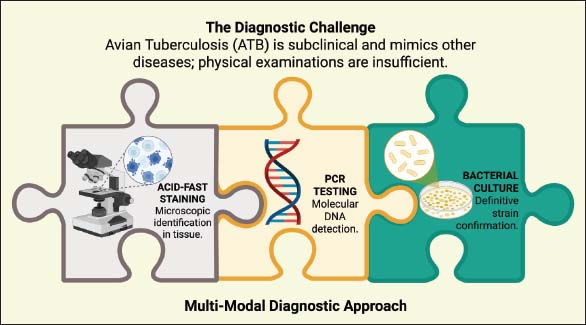
| How to Cite this Article |
| Pubmed Style Puspitasari Y, Khairullah AR, Raharjo HM, Tyasningsih W, Kurniasih DAA, Moses IB, Wardhani BWK, Ekawasti F, Pratama BP, Rehman S, Ahmad RZ, Abuzahra M, Utari F, Wibowo S, Ma'ruf IF, Prihandani SS. Current perspectives on avian tuberculosis in domestic and wild birds: Molecular diagnostics, genomic surveillance, and vaccination challenges. Open Vet. J.. 2026; 16(5): 2562-2580. doi:10.5455/OVJ.2026.v16.i5.1 Web Style Puspitasari Y, Khairullah AR, Raharjo HM, Tyasningsih W, Kurniasih DAA, Moses IB, Wardhani BWK, Ekawasti F, Pratama BP, Rehman S, Ahmad RZ, Abuzahra M, Utari F, Wibowo S, Ma'ruf IF, Prihandani SS. Current perspectives on avian tuberculosis in domestic and wild birds: Molecular diagnostics, genomic surveillance, and vaccination challenges. https://www.openveterinaryjournal.com/?mno=308127 [Access: June 26, 2026]. doi:10.5455/OVJ.2026.v16.i5.1 AMA (American Medical Association) Style Puspitasari Y, Khairullah AR, Raharjo HM, Tyasningsih W, Kurniasih DAA, Moses IB, Wardhani BWK, Ekawasti F, Pratama BP, Rehman S, Ahmad RZ, Abuzahra M, Utari F, Wibowo S, Ma'ruf IF, Prihandani SS. Current perspectives on avian tuberculosis in domestic and wild birds: Molecular diagnostics, genomic surveillance, and vaccination challenges. Open Vet. J.. 2026; 16(5): 2562-2580. doi:10.5455/OVJ.2026.v16.i5.1 Vancouver/ICMJE Style Puspitasari Y, Khairullah AR, Raharjo HM, Tyasningsih W, Kurniasih DAA, Moses IB, Wardhani BWK, Ekawasti F, Pratama BP, Rehman S, Ahmad RZ, Abuzahra M, Utari F, Wibowo S, Ma'ruf IF, Prihandani SS. Current perspectives on avian tuberculosis in domestic and wild birds: Molecular diagnostics, genomic surveillance, and vaccination challenges. Open Vet. J.. (2026), [cited June 26, 2026]; 16(5): 2562-2580. doi:10.5455/OVJ.2026.v16.i5.1 Harvard Style Puspitasari, Y., Khairullah, . A. R., Raharjo, . H. M., Tyasningsih, . W., Kurniasih, . D. A. A., Moses, . I. B., Wardhani, . B. W. K., Ekawasti, . F., Pratama, . B. P., Rehman, . S., Ahmad, . R. Z., Abuzahra, . M., Utari, . F., Wibowo, . S., Ma'ruf, . I. F. & Prihandani, . S. S. (2026) Current perspectives on avian tuberculosis in domestic and wild birds: Molecular diagnostics, genomic surveillance, and vaccination challenges. Open Vet. J., 16 (5), 2562-2580. doi:10.5455/OVJ.2026.v16.i5.1 Turabian Style Puspitasari, Yulianna, Aswin Rafif Khairullah, Hartanto Mulyo Raharjo, Wiwiek Tyasningsih, Dea Anita Ariani Kurniasih, Ikechukwu Benjamin Moses, Bantari Wisynu Kusuma Wardhani, Fitrine Ekawasti, Bima Putra Pratama, Saifur Rehman, Riza Zainuddin Ahmad, Mutasem Abuzahra, Fadhila Utari, Syahputra Wibowo, Ilma Fauziah Ma'ruf, and Sri Suryatmiati Prihandani. 2026. Current perspectives on avian tuberculosis in domestic and wild birds: Molecular diagnostics, genomic surveillance, and vaccination challenges. Open Veterinary Journal, 16 (5), 2562-2580. doi:10.5455/OVJ.2026.v16.i5.1 Chicago Style Puspitasari, Yulianna, Aswin Rafif Khairullah, Hartanto Mulyo Raharjo, Wiwiek Tyasningsih, Dea Anita Ariani Kurniasih, Ikechukwu Benjamin Moses, Bantari Wisynu Kusuma Wardhani, Fitrine Ekawasti, Bima Putra Pratama, Saifur Rehman, Riza Zainuddin Ahmad, Mutasem Abuzahra, Fadhila Utari, Syahputra Wibowo, Ilma Fauziah Ma'ruf, and Sri Suryatmiati Prihandani. "Current perspectives on avian tuberculosis in domestic and wild birds: Molecular diagnostics, genomic surveillance, and vaccination challenges." Open Veterinary Journal 16 (2026), 2562-2580. doi:10.5455/OVJ.2026.v16.i5.1 MLA (The Modern Language Association) Style Puspitasari, Yulianna, Aswin Rafif Khairullah, Hartanto Mulyo Raharjo, Wiwiek Tyasningsih, Dea Anita Ariani Kurniasih, Ikechukwu Benjamin Moses, Bantari Wisynu Kusuma Wardhani, Fitrine Ekawasti, Bima Putra Pratama, Saifur Rehman, Riza Zainuddin Ahmad, Mutasem Abuzahra, Fadhila Utari, Syahputra Wibowo, Ilma Fauziah Ma'ruf, and Sri Suryatmiati Prihandani. "Current perspectives on avian tuberculosis in domestic and wild birds: Molecular diagnostics, genomic surveillance, and vaccination challenges." Open Veterinary Journal 16.5 (2026), 2562-2580. Print. doi:10.5455/OVJ.2026.v16.i5.1 APA (American Psychological Association) Style Puspitasari, Y., Khairullah, . A. R., Raharjo, . H. M., Tyasningsih, . W., Kurniasih, . D. A. A., Moses, . I. B., Wardhani, . B. W. K., Ekawasti, . F., Pratama, . B. P., Rehman, . S., Ahmad, . R. Z., Abuzahra, . M., Utari, . F., Wibowo, . S., Ma'ruf, . I. F. & Prihandani, . S. S. (2026) Current perspectives on avian tuberculosis in domestic and wild birds: Molecular diagnostics, genomic surveillance, and vaccination challenges. Open Veterinary Journal, 16 (5), 2562-2580. doi:10.5455/OVJ.2026.v16.i5.1 |

