




| Research Article | ||
Open Vet. J.. 2026; 16(5): 2987-3007 Open Veterinary Journal, (2026), Vol. 16(5): 2987-3007 Research Article Decoding antidengue serotype-3 (DENV-3) mechanism of natural glycosides from Alpinia purpurata: Molecular docking and in vitro insights into viral entry, cytokine storm modulation, and endogenous antioxidant defense activationMochammad Aqilah Herdiansyah1, Win Darmanto2*, Dwi Winarni2, Aulia Umi Rohmatika3, Rr Aulia Rahmawati Kusuma Putri1, Raden Joko Kuncoroningrat Susilo4, Teguh Hari Sucipto5, Putu Angga Wiradana6, Farach Khanifah7 and Ernawati Ernawati81Doctoral Program of Mathematics and Natural Sciences, Faculty of Science and Technology, Universitas Airlangga, Surabaya, Indonesia 2Department of Biology, Faculty of Science and Technology, Universitas Airlangga, Surabaya, Indonesia 3Faculty of Medicine, Universitas Pembangunan Nasional Veteran Jawa Timur, Surabaya, Indonesia 4Nanotechnology Engineering, Faculty of Advance Technology and Multidiscipline, Universitas Airlangga, Surabaya, Indonesia 5Dengue Study Group, Institute of Tropical Disease, Universitas Airlangga, Surabaya, Indonesia 6Study Program of Biology, Faculty of Health and Science, Universitas Dhyana Pura, Bali, Indonesia 7Teknologi Laboratorium Medis, Institut Teknologi Sains dan Kesehatan Insan Cendekia Media Jombang, Jombang, Indonesia 8Faculty of Health, Universitas Muhammadiyah Gresik, Kebomas, Indonesia *Corresponding Author: Win Darmanto. Department of Biology, Faculty of Science and Technology, Universitas Airlangga, Surabaya, Indonesia. Email: windarmanto [at] fst.unair.ac.id Submitted: 30/12/2025 Revised: 19/03/2026 Accepted: 31/03/2026 Published: 31/05/2026 © 2025 Open Veterinary Journal
AbstractBackground: Dengue virus serotype-3 (DENV-3) infection remains a clinical problem for which no specific antiviral agent has yet been identified. The rhizome of Alpinia purpurata (red galangal) is rich in natural glycosides with antioxidant and anti-inflammatory properties. However, studies related to antidengue from A. purpurata using in vitro and in silico methods have not been widely conducted. Aim: This study aims to describe the anti-dengue mechanism of natural glycosides from A. purpurata rhizomes through their antiviral ability in vitro in Vero E6 cell cultures induced by DENV-3 and molecular docking to the DC-SIGN (CD209), CD300a, IL-1β, and SOD receptors. Methods: In vitro quantification of the viral replication inhibition value was performed using dengue virus antibodies (4G2) through ELISA. Meanwhile, molecular docking was performed to assess the binding affinities and complex interactions. Molecular dynamics simulations confirmed the stability of the docking complexes within the root mean square fluctuation value. Further screening was conducted using the pkCSM webserver to obtain the ADMET property values and the ProTox 3.0 webserver to obtain the toxicity prediction values. Results: In vitro DENV-3-induced Vero E6 cell cultures induced with DENV-3 showed a viral replication inhibition value of 105.8107 µg/mL. Molecular docking experiments revealed that quercetin-3-O-rutinoside has the best potential to inhibit virus binding to DC-SIGN receptors (−7.8 kcal/mol), balance immune signals through CD300a regulation (−7.1 kcal/mol), suppress the production of IL-1β during cytokine storm (−7.2 kcal/mol), and activate endogenous antioxidant superoxide dismutase (−7.0 kcal/mol) compared with quercetin control. The validation of docking results through molecular dynamics proved that the quercetin-3-O-rutinoside complex has an RMSF value < 3 Å, which means that the complex tends to be stable. ADMET characteristics and toxicity also indicate that quercetin-3-O-rutinoside is safe for cells. Conclusion: This study reports the first evidence of the potential of natural glycosides from A. purpurata as antidengue agents in vitro and in silico through viral entry inhibition, cytokine storm modulation, and endogenous antioxidant activation to reduce free radicals in cells during infection. These findings support the potential of A. purpurata as an antiviral, anti-inflammatory, and antioxidant agent. Keywords: Alpinia purpurata; Antidengue; Glycosides; Multiprotein Target. IntroductionDengue fever is a viral infectious disease caused by the dengue virus (DENV), which is transmitted by the mosquito vector of Aedes aegypti. This infectious disease remains a global health problem because no specific effective antiviral drug or vaccine that fully protects cells from viral infection exists (Islam et al., 2025). Current therapeutic strategies focus on discovering natural compounds that can inhibit the early stages of infection and modulate the immune response and intracellular oxidative stress. Based on the latest epidemiological and molecular analyses, DENV-1, DENV-2, and DENV-3 are the most prevalent serotypes, and shifts in serotype dominance are often associated with spikes in cases and increased disease severity (Harapan et al., 2019). In recent years, DENV-3 has reemerged as the dominant serotype in various regions of Indonesia, such as Yogyakarta and East Java, with genetic characteristics indicating a shift in genotype and increased virulence (Utama et al., 2019; Aryati et al., 2020). In addition, the dominance of DENV-3 in Southeast Asia has also been found in Myanmar and Bangladesh, which have reported the emergence of new genotypes (types I and III) associated with an increase in severe cases and expansion of the transmission area (Soe et al., 2021; Titir et al., 2021). The interaction between the envelope protein and the dendritic cell-specific intracellular adhesion molecule-3-grabbing non-integrin (DC-SIGN/CD209) receptor is one of the main mechanisms of DENV-3 invasion. This protein is expressed by the dendritic cells of the body and indirectly helps the virus enter the host body. This interaction facilitates virus attachment and internalization into the host cell (Singh et al., 2021). In addition to DC-SIGN, the CD300a receptor regulates inflammatory signals and balances protective immune responses and excessive inflammation. In the later stages of DENV-3 infection, the excessive release of proinflammatory cytokines such as IL-1β in patients infected with DENV can trigger a “cytokine storm” mechanism that contributes to severe pathogenesis, such as dengue hemorrhagic fever and dengue shock syndrome (Alam et al., 2020). Therefore, modulation of IL-1β expression is needed to suppress inflammation in the body. Additionally, activation of the endogenous antioxidant protein synthesis pathway, such as superoxide dismutase (SOD), may also be key in reducing tissue damage caused by oxidative stress during viral infection. Plants from the Alpinia genus, particularly Alpinia purpurata, are endemic in Southeast Asia (Kong et al., 2016). These plants are rich in natural glycoside compounds with broad biological activities, including antioxidant, anti-inflammatory, and antiviral properties (Aguinaldo et al., 2010). Glycoside compounds can interact with C-type lectin receptors, such as DC-SIGN, through specific affinity for mannose or galactose residues on their surface (Tanaka et al., 2017). This binding mechanism can inhibit virus attachment and modulate immune signals activated by DC-SIGN and CD300a receptors. In silico molecular docking approaches have been widely used to identify and predict interactions between natural compounds and DENV target proteins, such as NS3 protease, NS5 methyltransferase, and host immune receptors (Imran et al., 2025). Several natural phytochemical compounds, such as α-Mangostin from Garcinia mangostana, have been shown to exhibit antiviral activity against DENV-2 through a multiprotein virus inhibition mechanism both in silico and in vitro (Panda et al., 2021). Based on this, this study aims to describe the anti-dengue mechanism of natural glycosides from A. purpurata rhizomes through a molecular docking approach to the DC-SIGN (CD209), CD300a, IL-1β, and SOD receptors, as well as to validate their antiviral ability in vitro against DENV-3-induced Vero E6 cell cultures induced by DENV-3. This study is expected to provide a scientific basis for the development of new natural drug candidates for multitarget and immunomodulatory dengue therapy. Materials and MethodsSample collection and extract preparationAlpinia purpurata rhizome samples were collected from Tanjung Market, Mojokerto, East Java, Indonesia (latitude: 7°28'1“S; longitude: 112°26'12”E). Preparation of the extract began by cutting the A. purpurata rhizomes into small pieces and drying them in the sun for 14 days. Drying was performed out by sifting the material once a day to avoid fungi contamination. Next, the material was macerated with 96% ethanol at a ratio of 1:3. Maceration was performed for 7 days with the aim of extracting the secondary metabolites in the rhizome by stirring occasionally. The ethanol extract is filtered, and the residue is re-extracted using the same procedure. Then, evaporation using a rotary evaporator is performed at a temperature of 40℃–60℃, a vacuum pressure of 100–200 mbar, and a rotation speed of 80–120 rpm. The extract was scraped and stored in test bottles. Preparation of cell cultureThe Vero E6 cell culture collection (ATCC® CRL-1586™) was obtained from the Institute of Tropical Diseases, Airlangga University, East Java, Indonesia. Cells were maintained in Dulbecco’s Modified Eagle Medium (DMEM) supplemented with 10% heat-inactivated fetal bovine serum (FBS) and 1% penicillin-streptomycin and incubated at 37℃ with 5% CO2. Monolayer cell cultures that had reached confluence (80%) were trypsinized using 0.25% trypsin-EDTA, and their viability was calculated using a hemocytometer. The cells were then transplanted into 96-well plates. Next, cells were allowed to adhere for 24 hours before DENV-3 inoculation at a multiplicity of infection (MOI) optimized between 0.1 and 1.0, following established protocols for dengue antiviral screening in Vero lineages (Abdul Ahmad et al., 2017; Low et al., 2021). Antiviral assay to determine the percent inhibition of virus replication valueQuantification of the viral replication inhibition value of the extract and quercetin control was performed using dengue virus antibodies (4G2) through an ELISA. Fixation in the well was fixed using methanol. Acetone and blocking buffer (2% skim milk, 0.05% Tween-20 PBS) were added to each well. After 30 min, the primary antibody, 4G2 antibody, was added and incubated for 1 hour at 37°C for 1 hour. Washing was performed four times using washing buffer (0.01% Tween-20 PBS). The secondary antibody, goat anti-mouse IgG HRP, was added and incubated for 1 hour at 37°C for 1 hour. The washing was repeated four times using a washing buffer. Next, TMB substrate was added and incubated for 10 min until a color change occurred. Subsequently, a 2 N H2SO4 solution was added to stop the reaction and measured using a microplate reader at 450 nm (Koishi et al., 2012). The EC50 value represents the inhibition of virus replication performance due to treatment (Sucipto et al., 2017). In silico studyHardwareThe in silico method was performed using a Dell Vostro 14 3000 laptop with an AMD Ryzen 5 3500 U processor. The operating system used was Windows 10 Ultimate 64-bit with Radeon Vega Mobile Gfx at 2.10 GHz. Ligand preparationThe test ligands used were natural glycosides found in A. purpurata. The names of each test ligand were obtained from previous relevant metabolite screening studies using A. purpurata. Some of the test ligands used in this study were sitosteryl-3-O-6-palmitoyl-β-D-glucoside (CID: 70699351) (Villaflores et al., 2010 ), quercetin-3-O-rutinoside (CID: 5280805) (Victório et al., 2019; Suzery et al., 2019), kaempferol-3-O-glucuronide (CID: 74315888) (Victório et al., 2019), vanillic acid glucoside (CID: 14132336) (Begum et al., 2019), and p-coumaric acid glucoside (CID: 9840292) (Begum et al., 2019). Quercetin control (CID: 5280343) was used as a reference. All canonical SMILES, PDB ID, and 3D structure information related to the test compounds were obtained from the PubChem webserver (https://pubchem.ncbi.nlm.nih.gov/). Protein preparationThe test protein was prepared by downloading the receptor file from the Uniprot webserver (https://www.uniprot.org/). The test receptors used were DC-SIGN (CD209) (Uniprot ID: Q9NNX6), CD300a (Uniprot ID: Q9UGN4), IL-1β (Uniprot ID: P01584), and superoxide dismutase (SOD) (Uniprot ID: P00441) (Table 1). Protein sterilization was performed using PyMol v1.74 software (Schrödinger, LLC). The purpose of this sterilization was to remove water molecules and non-essential ligands that were still present on the test receptors (Siti Rahayu et al., 2025). Table 1. List of receptors used in the molecular docking experiments.
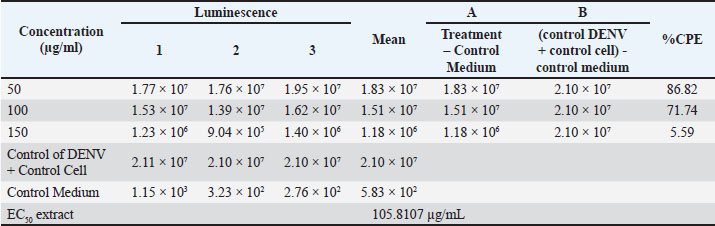
Molecular docking experiment and antiviral potential screeningMolecular docking experiments were conducted using PyRx v.0.8 software (SourceForge Headquarters, San Diego, CA) to obtain binding affinity (kcal/mol) and root mean square deviation (RMSD) (Å) values between the test ligand and target receptor. The binding affinity value refers to the energy generated between the ligand-protein complex. The more negative the binding affinity value, the higher the potential of the compound to trigger an anti-dengue bioactivity response (Alifiansyah et al., 2024; Herdiansyah et al., 2024; Vora et al., 2020). The PASS Online webserver (https://way2drug.com/PassOnline/) was used to screen the antiviral potential of each compound to determine the probable of active (Pa) and probable of inactive (Pi) values. A compound is considered to have antiviral potential if it has a Pa>Pi value. Compounds with the best antiviral bioactivity potential based on binding affinity, RMSD, and probable of active values will be selected for further screening. Post-screening of selected potential compounds based on the absorption, distribution, metabolism, excretion, and toxicity (ADMET) properties and specific toxicityScreening of pharmacokinetic parameters of glycoside compounds with the best antiviral bioactivity potential from molecular docking results was performed using the pkCSM webserver (https://biosig.lab.uq.edu.au/pkcsm/). The pkCSM webserver analyzes the relationship between the molecular structure presented in the form of SMILES code of the compound and the pharmacokinetic behavior of the drug in the body, which can be used as an evaluation of the safety of the compound when in the body. There are five major groups of parameters, namely absorption, distribution, metabolism, excretion, and toxicity, which are analyzed in this ADMET screening, with each parameter group having its own sub-parameters (Fatima et al., 2020; Klimoszek et al., 2024; Banerjee et al., 2024). Furthermore, the specific toxicity profile of glycoside compounds with the best bioactivity potential was predicted using the ProTox 3.0 webserver (https://tox.charite.de/protox3/). ProTox integrates molecular similarity, compound pharmacophore models, fragments, and machine learning models to predict the toxicity of a compound, including acute toxicity (LD50), hepatotoxicity, carcinogenicity, mutagenicity, immunotoxicity, cytotoxicity, and organ-specific toxicities (Banerjee et al., 2024). Molecular dynamics simulation for validation of docking resultsThe molecular docking experiment results were validated using the CABS-flex 3.0 webserver (https://lcbio.pl/cabsflex3/). The results with the best binding affinity values were used for validation to observe the properties of the drug when attached to the protein ring. The molecular docking results were validated using several parameters, such as protein rigidity, restraint settings, C-alpha restraints weight, side-chain restraint weight, temperature range, trajectory, number of cycles, and RNG seed in nanoseconds (ns) (Shivanika S Deepak et al., 2022). Statistical analysisThe data were statistically analyzed using SPSS 21 software (SPSS Inc., Chicago, IL). Data were analyzed using t-test, and differences between mean values were considered significant at p < 0.05. Meanwhile, for p > 0.05, non-significant differences were considered between extract and control, and accepting H0. Ethical approvalNot needed for this study. ResultsIn vitro antiviral assay results of inhibition of virus replicationThe ethanol extract of A. purpurata inhibits DENV-3 replication in a concentration-dependent manner. As shown in Table 2, at a concentration of 50 µg/ml, the extract provided 86.82% inhibition of the cytopathic effect (CPE), which decreased to 71.74% at 100 µg/ml and 5.59% at 150 µg/ml. The EC50 value of 105.81 µg/ml indicates moderate effectiveness against DENV-3, which is comparable to that of the quercetin control (EC50=92.69 µg/ml). The natural glycoside content in A. purpurata extract can be a natural antiviral source against DENV-3. Table 2. Antiviral assay of the extract against DENV-3 infection.
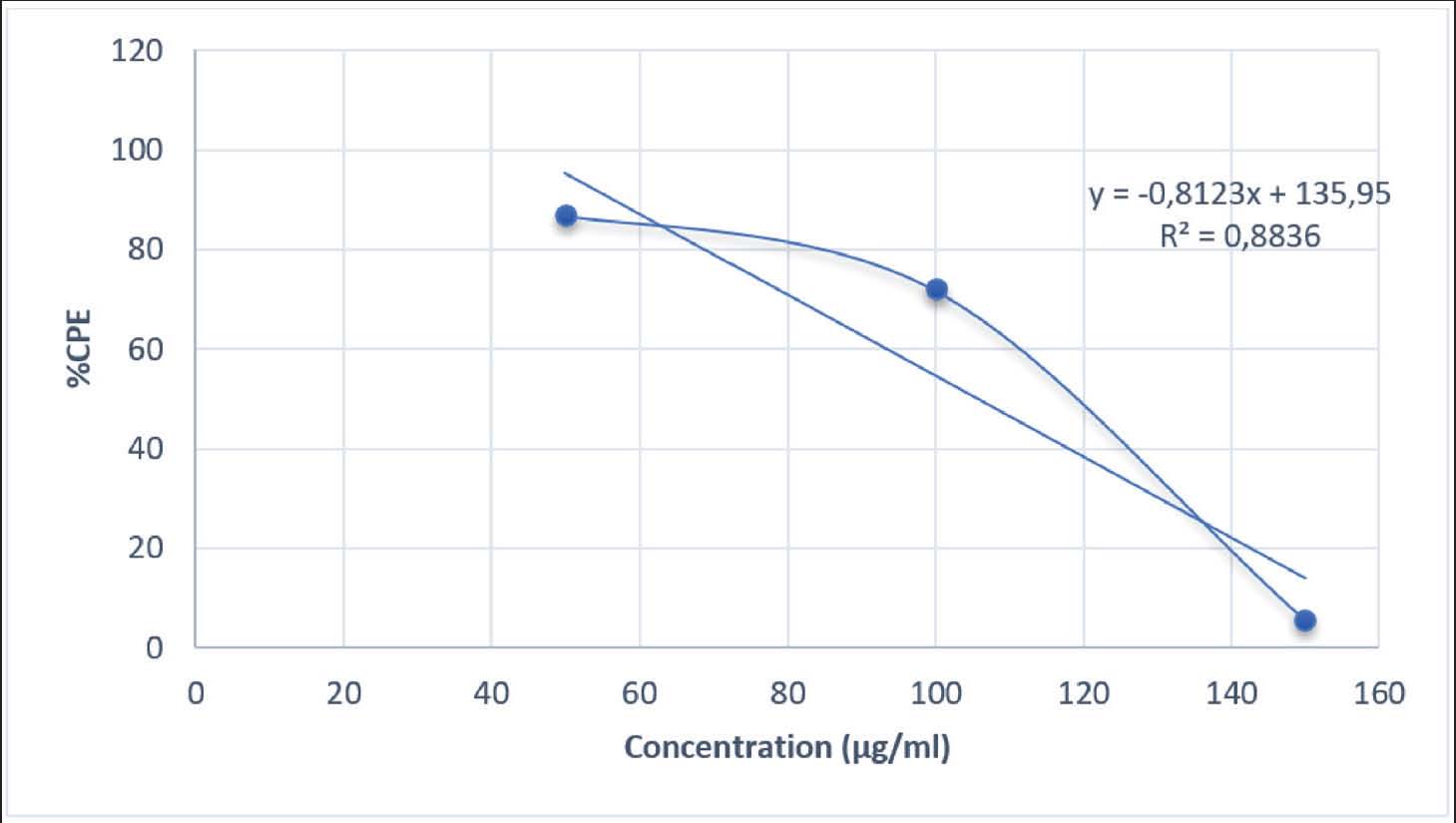
The highest inhibitory effect at a concentration of 50 µg/ml in this study indicates that at low doses, the active compounds in the extract are still within the non-toxic range for host cells, thereby effectively inhibiting viral replication. The decrease in activity at higher concentrations may be attributed to mild cytotoxic effects or oxidative stress, which reduces cell viability and viral inhibition effectiveness. The recorded luminescence values also support this result, as higher extract concentrations corresponded to lower luminescence signals due to reduced viral infectivity. However, at a concentration of 150 µg/mL, the luminescence value dropped sharply, possibly due to cellular metabolic disturbances caused by an excess of active compounds in the extract. The inhibitory activity in Table 2 likely involves a synergistic mechanism between virus attachment inhibition and intracellular immune response modification. This mechanism can also be observed in the natural compounds α-mangostin and artesunic acid, which suppress DENV replication through multiprotein interaction in the early and late phases of the virus life cycle. The downward trend in CPE percentage in Fig. 1 is consistent with the increasing concentration of A. purpurata extract against DENV-3 infection in Vero E6 cells. This phenomenon is commonly found in plant extracts rich in polyphenol and glycoside secondary metabolites. At low doses, active compounds tend to work optimally in suppressing viral replication through a mechanism that inhibits virus attachment to cell receptors without disrupting cell viability. However, at higher doses, the accumulation of phenolic compounds can increase oxidative stress and disrupt the integrity of the host cell membrane. This can reduce overall antiviral effectiveness (Rosmalena et al., 2019).
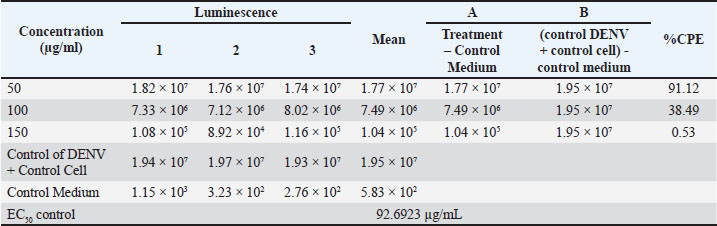
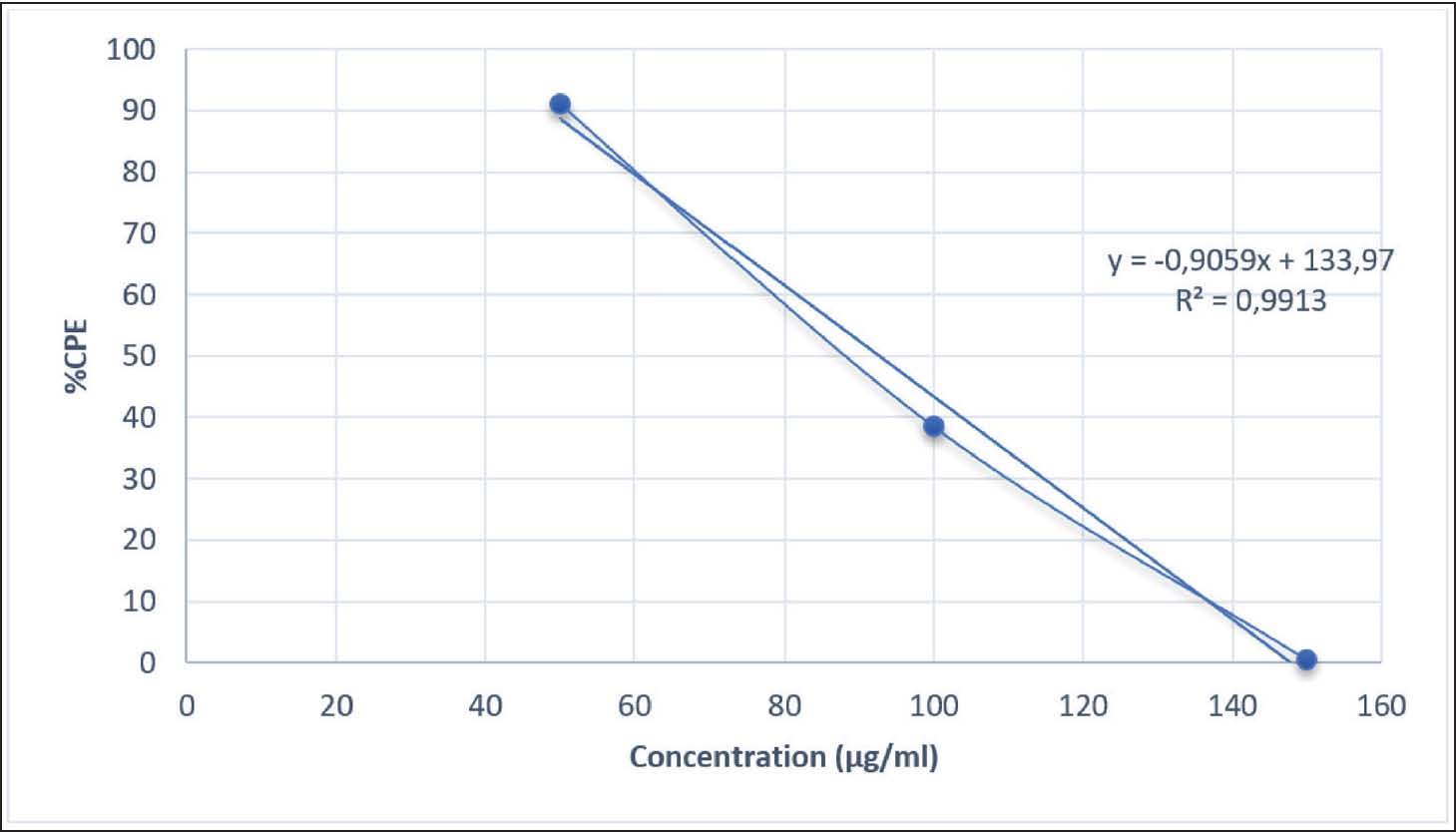
Fig. 1. Cytopathic effect percentage of extracts that inhibit DENV-3 infection in the cell line. At low to moderate doses, the active compounds suppress viral replication by inhibiting viral attachment to cell receptors without affecting host cell viability. However, excessive accumulation of phenolic compounds at high doses may induce mild oxidative stress and disrupt cell membrane integrity, which in turn can reduce the antiviral effectiveness of the extract. Therefore, the decrease in the CPE curve at higher concentrations may also reflect a shift in the antiviral mechanism of action—from early-stage (penetration) inhibition to viral replication suppression within Vero E6 cells. A comparison between the antiviral test results of A. purpurata extract (Table 2, Fig. 1) and quercetin control (Table 3, Fig. 2) showed a similar biological response pattern, but with different levels of effectiveness. At a concentration of 50 µg/mL, A. purpurata extract reduced cytopathic effects by 86.82%, whereas quercetin reduced cytopathic effects by 91.12% at the same concentration. Although this difference is relatively small, the EC50 value of the extract is slightly lower than that of quercetin, indicating that the antiviral bioactivity of the extract is similar to that of pure flavonoid compounds. Thus, the glycoside compounds in the A. purpurata extract have competitive antiviral potential despite originating from a mixture of natural secondary metabolites. Table 3. Antiviral assay of quercetin control against DENV-3 infection.
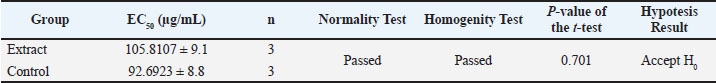
Fig. 2. Cytopathic effect percentage of the control that inhibited DENV-3 infection in the cell line. The downward trend in CPE percentage in both samples also showed a dose-dependent biphasic response pattern, in which antiviral efficacy increased at low to medium doses but decreased at high doses (150 µg/mL). Although the control had a better EC50 value than the test extract, the A. purpurata extract showed the potential advantage of a multicomponent synergistic effect. Natural glycoside components, such as quercetin-3-O-rutinoside, kaempferol-3-O-glucuronide, and sitosteryl-3-O-6-palmitoyl-β-D-glucoside, can act through various targets, including inhibition of virus binding to DC-SIGN/CD209 receptors and modulation of inflammatory pathways via IL-1β. This multitarget protein approach makes the extract more relevant than single compounds for natural drug development. Statistical test results showed no significant difference between the extract test group and the control group (p > 0.05). This value is greater than the significance threshold of 0.05 (Table 4); therefore, the anti-dengue activity of A. purpurata extract against DENV-3 is comparable to quercetin. This finding is consistent with several previous studies reporting that flavonoids and polyphenols from plants have similar antiviral mechanisms by inhibiting dengue virus replication and attachment to host cells. Interestingly, quercetin and fisetin can reduce DENV-2 and DENV-3 virus titers and suppress proinflammatory cytokine secretion during infection (Jasso-Miranda et al., 2019). Therefore, further predictions using molecular docking and molecular dynamics experiments are needed to identify potential single compounds from A. purpurata extract, particularly from the abundant glycoside group in the plant. Table 4. Analysis of antiviral assay results.
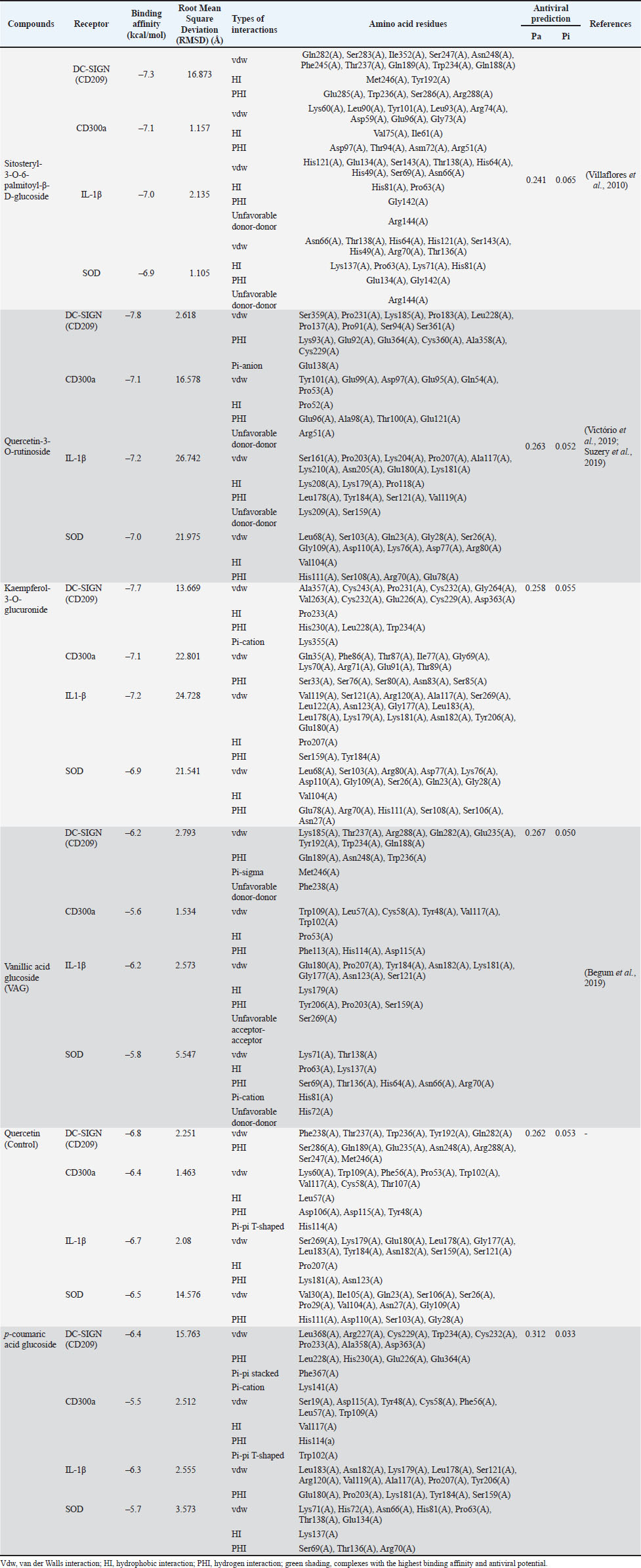
Molecular docking of glycosides and test receptorsThe in silico analysis in Table 5 shows that the main glycoside compounds from A. purpurata have a strong binding affinity to host cell receptors that play a role in DENV infection and immune regulation. The highest binding energy, greater than that of the control, was found in the compound quercetin-3-O-rutinoside (DC-SIGN=−7.8 kcal/mol; CD300a=−7.1 kcal/mol; IL-1β=−7.2 kcal/mol; SOD=−7.0 kcal/mol). This pattern indicates that A. purpurata glycosides have the potential to inhibit virus binding and stabilize the cellular immune response. Table 5. Molecular docking experiment and antiviral prediction results for each glycoside-receptor complexes.
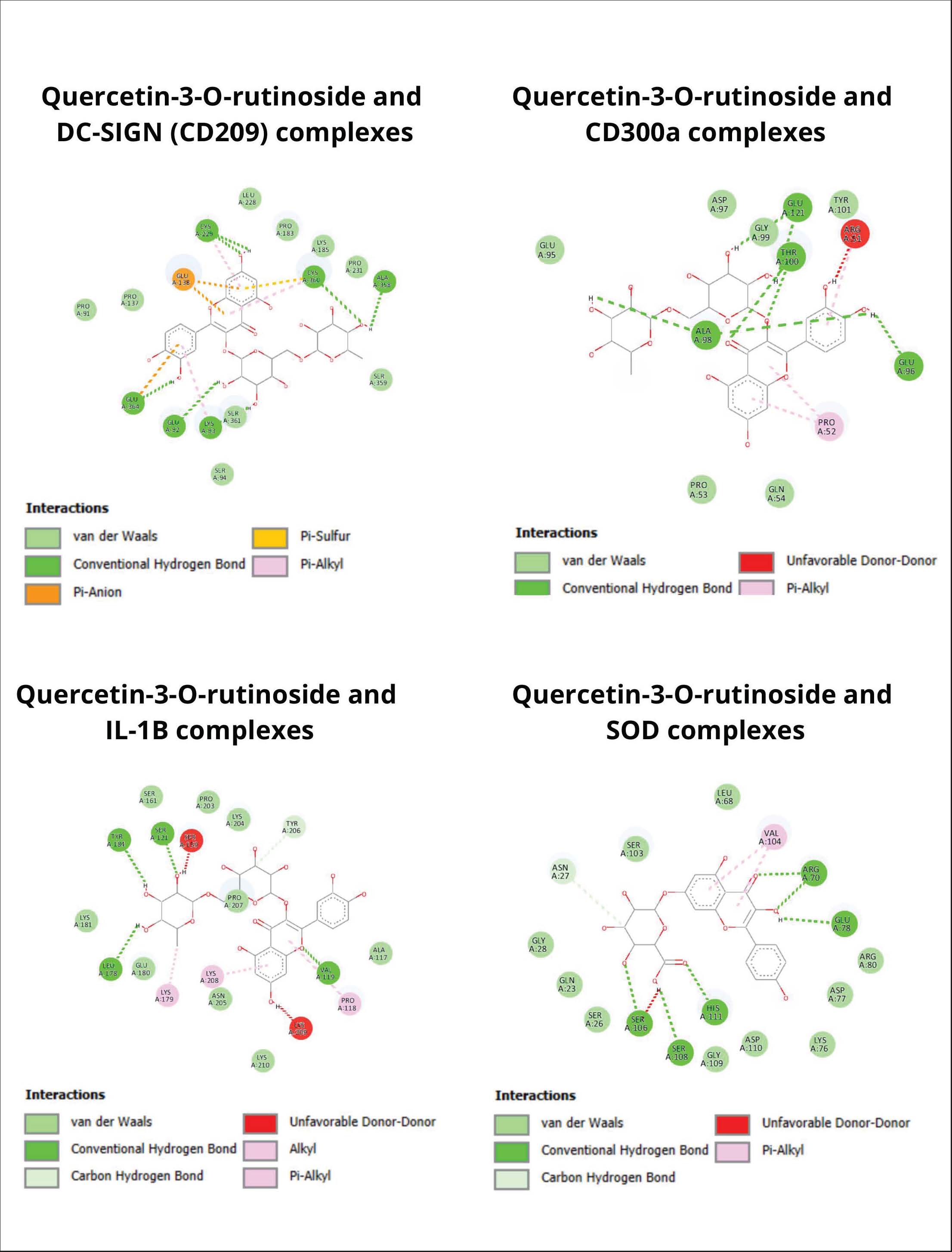
In Table 5, strong interactions were detected through the amino acid residues Gln282(A), Ser283(A), Ile352(A), and Trp236(A), which contribute to the formation of hydrogen bonds and Van der Waals forces. This interaction pattern indicates that the glycoside can inhibit the attachment of viral particles to the DC-SIGN receptor, which serves as the main entry point for DENV-3 into host cells. The glycoside compound quercetin-3-O-rutinoside inhibited DC-SIGN and showed a significantly stronger binding pattern than the positive control (−6.8 kcal/mol), suggesting that the compound from the extract possesses a better inhibitory capacity in preventing viral entry into the host. In addition, glycoside compounds were found to interact with the residues Lys60(A), Leu90(A), Arg74(A), and Asp97(A) through hydrogen bonds and Van der Waals forces on the CD300a receptor. This pattern illustrates the potential of glycosides to modulate immune activity, as CD300a functions as an immune signal regulatory receptor that can be exploited by DENV-3 to suppress phagocytosis. The binding of glycosides to these amino acid residues is presumed to inhibit the CD300a-mediated inhibitory signals, thereby assisting in the restoration of the immune response following DENV-3 infection. Significant interactions were also detected between the ligand and the IL-1β receptor, in which the residues His121(A), Glu134(A), and Gly142(A) were directly involved in forming strong hydrogen bonds and Van der Waals forces. These interactions contribute to the suppression of signaling pathway activation that leads to the production of the proinflammatory cytokine IL-1β during cytokine storm conditions within the cell. This pattern indicates that the glycoside compounds of A. purpurata function not only as anti-dengue agents but also as natural immunomodulators that reduce the secretion of proinflammatory cytokines during infection. Meanwhile, hydrogen bonds were naturally formed between the ligand and the residues His111(A), Asp110(A), and Ser103(A) on the SOD enzyme, which play a role in maintaining the stability of the enzyme’s active site. The attachment of the tested compound to these amino acid residues may help the cell activate SOD enzyme secretion, thereby reducing free oxidants in the body when cells experience stress due to dengue infection. In addition, this interaction demonstrates the potential of glycosides to enhance cellular antioxidant defenses by preserving SOD enzyme function from oxidative stress-induced damage. The stability of this complex indicates that the active compounds of A. purpurata may support the cell’s natural protective mechanisms against increased free radicals during infection. Overall, Figure 3 supports the interactions shown in Table 5, which describes the synergistic antiviral-immunomodulator-antioxidant mechanism of A. purpurata extract. Glycoside compounds not only inhibited virus binding to DC-SIGN receptors but also suppressed the activation of proinflammatory cytokine IL-1β, balance immune signaling through the regulation of the CD300a protein, and strengthened the oxidative defense system through the superoxide dismutase enzyme. This combination of activities indicates that A. purpurata works through multireceptor modulation, which not only suppresses viral replication but also reduces pathogenesis in severe dengue infection due to immune hyperactivation.
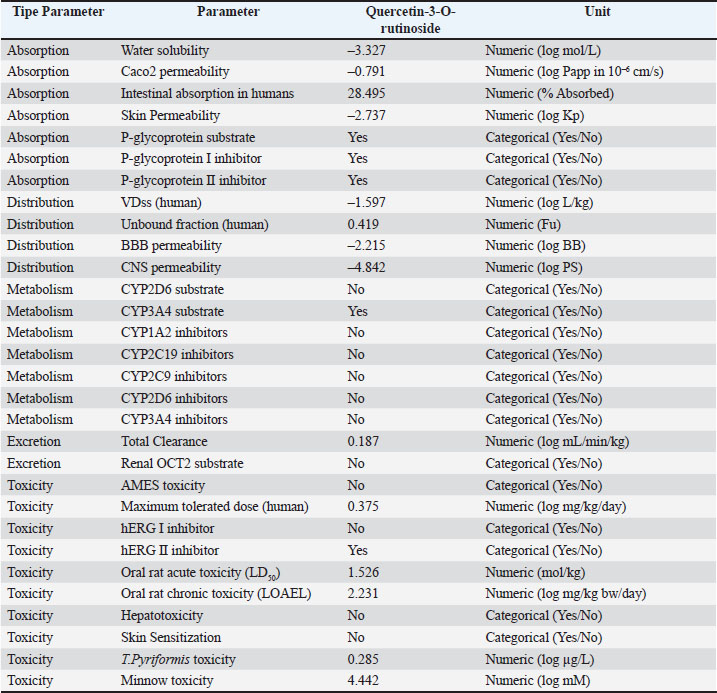
Fig. 3. Interaction complex visualization of quercetin-3-O-rutinoside as the complex with the highest binding affinity. ADMET properties and specific toxicity of quercetin-3-O-rutinosideThe aim of the ADMET analysis was to assess the pharmacokinetic potential and safety of the main active compound resulting from docking with the highest binding affinity value, quercetin-3-O-rutinoside from A. purpurata (Mvondo et al., 2021). This parameter is important for assessing the suitability of the compound as a drug candidate before conducting in vivo or clinical trials. The results in Table 6 show that the quercetin-3-O-rutinoside compound has moderate solubility (log S=−3.327), indicating a moderate ability to dissolve in cells. However, the Caco2 permeability value (−0.791 log Papp) and intestinal absorption of 28.49% indicate that the absorption through human milk is relatively lower due to the large glycoside structure and high polarity. This compound is also detected as a p-glycoprotein substrate and p-glycoprotein I and II inhibitor, which means that an efflux mechanism that limits oral bioavailability may occur (Fatima et al., 2020; Klimoszek et al., 2024). Table 6. ADMET properties of quercetin-3-O-rutinoside.
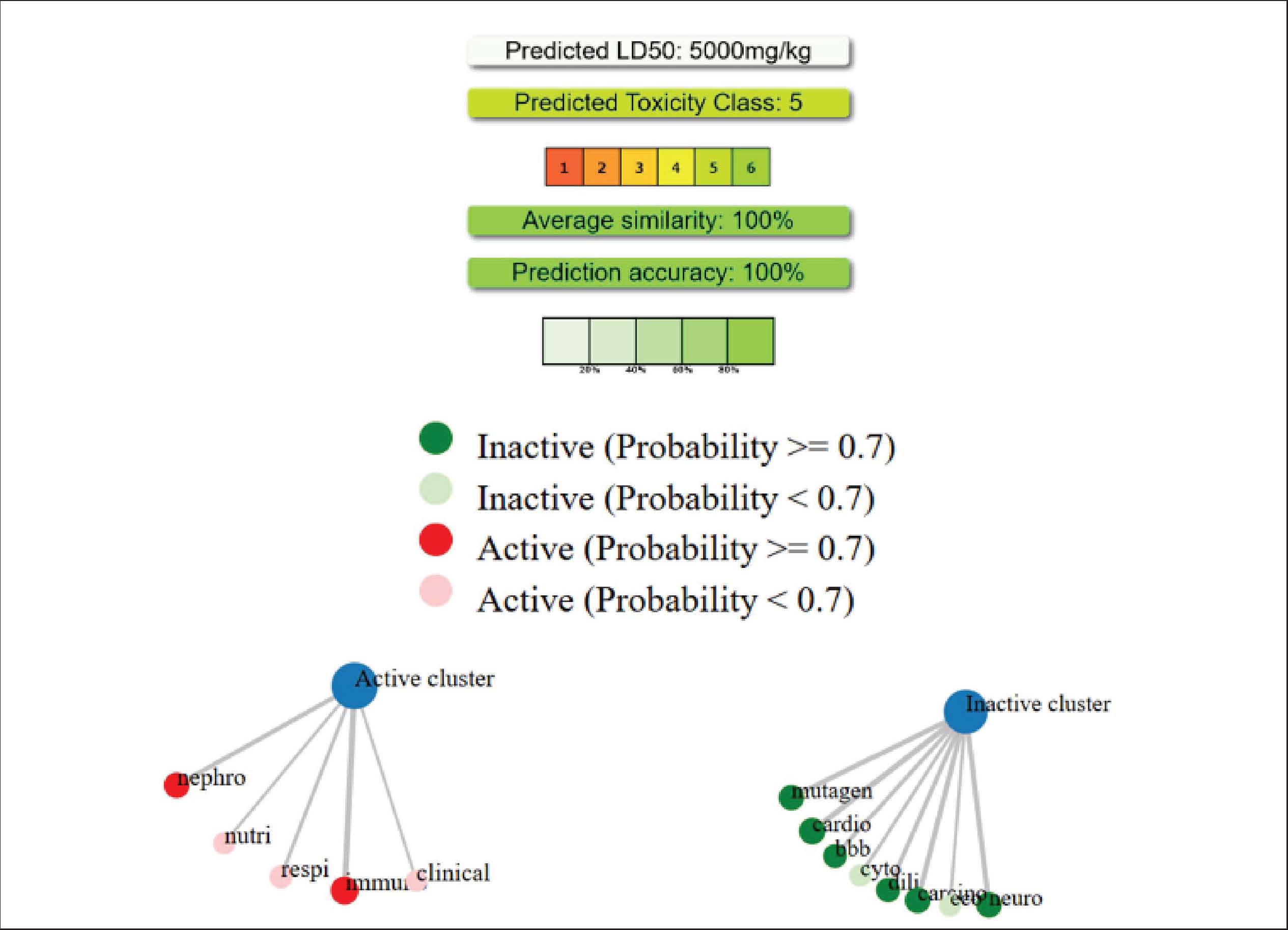
The volume of distribution (VDss) value of −1.597 log L/kg indicates low distribution to peripheral tissues, while the unbound fraction of 0.419 indicates that nearly half of the compound is in free form in plasma, allowing for effective pharmacological interactions. The BBB and CNS permeability values also indicate that this compound is suitable for preventing neurotoxicity or CNS disorders. The compound quercetin-3-O-rutinoside is not a CYP2D6 substrate but a CYP3A4 substrate in terms of distribution parameters. This indicates the potential for major metabolism via the cytochrome P450 pathway in the liver. The absence of inhibitory activity against major cytochrome P450 enzymes indicates a low risk of drug interactions. This pattern is similar to the ADMET profile of other natural flavonoid compounds, such as kaempferol-3-glucuronide, which is metabolized via CYP3A4 without causing hepatotoxicity (Shivanika S Deepak et al., 2022). In terms of excretion parameters, the total clearance value of 0.187 log ml/min/kg indicates slower excretion and supports a moderate half-life. Quercetin-3-O-rutinoside is not a renal OCT2 substrate, indicating that the main elimination route is hepatic or biliary (Herdiansyah et al., 2024). In addition, based on pkCSM predictions, quercetin-3-O-rutinoside does not exhibit genetic toxicity (Ames test negative), is not hepatotoxic, and does not cause skin sensitization. As shown in Figure 4, no hepatotoxic potential was found. This indicates that the quercetin-3-O-rutinoside compound cannot induce hepatocyte damage. This result is consistent with the pkCSM prediction for hepatotoxicity parameters. No mutagenic or carcinogenic potential was found, indicating that the compound does not contain genotoxic chemical fragments, such as long conjugated aromatic rings or active electrophilic substituents. Flavonoids with glycoside groups stabilize DNA through noncovalent interactions that protect against gene mutations. The absence of immunotoxicity indicates that quercetin-3-O-rutinoside has immunomodulatory activity and the potential to suppress proinflammatory cytokine production and normalize CD300a receptor expression. The LD50 value of 5000 mg/kg in the compound’s toxicity class 5 indicates a low tendency for acute toxicity.
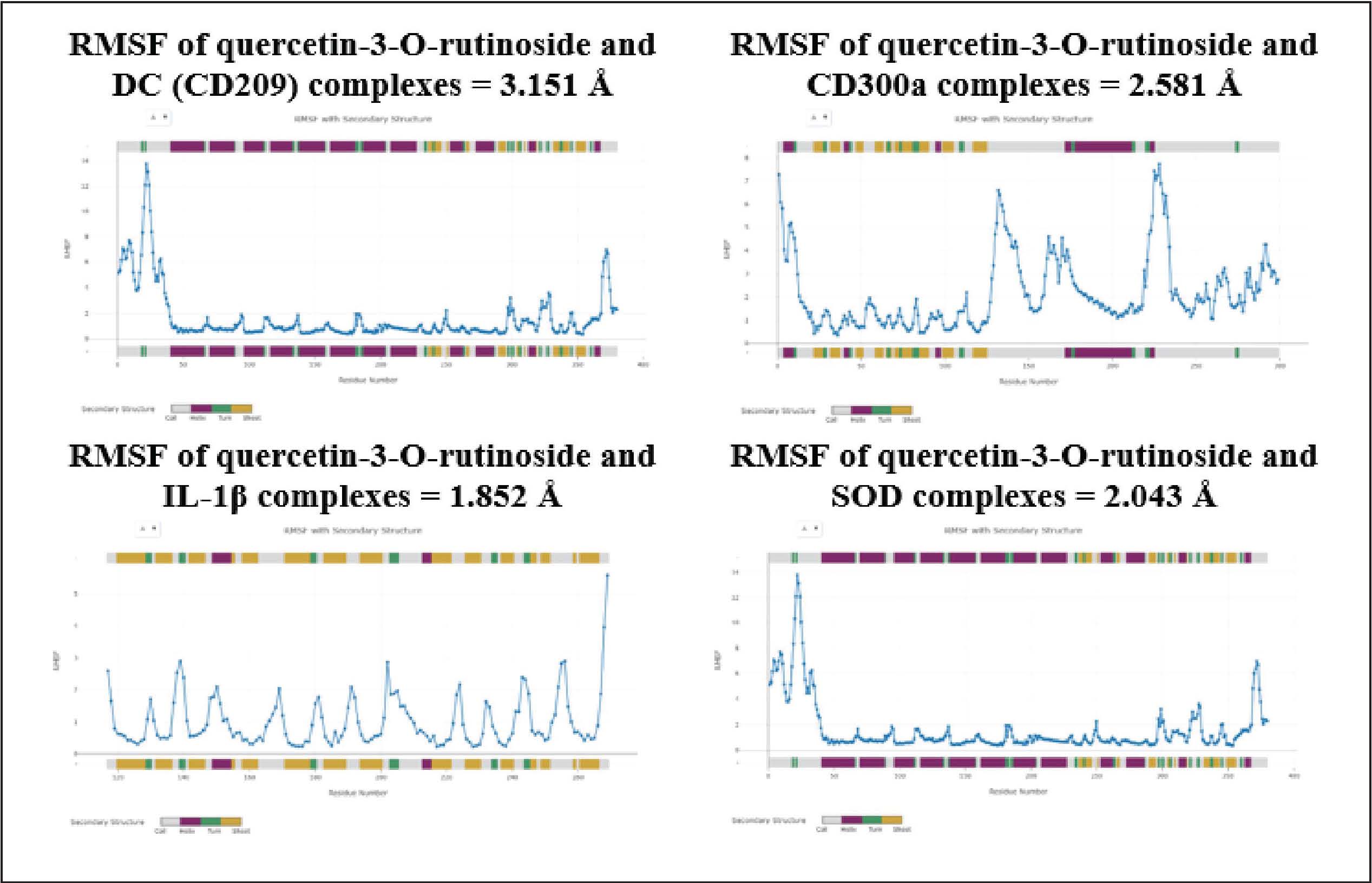
Fig. 4. Results of the toxicity analysis of quercetin-3-O-rutinoside on specific organs. Molecular dynamics results as validation of the molecular docking methodThe results of molecular docking validation through molecular dynamics show that the binding between quercetin-3-O-rutinoside and CD300a, IL-1β, and SOD is stable. However, the complex between quercetin-3-O-rutinoside and DC (CD209) has an RMSF value > 3 Å. The instability of the bond between the receptor and the ligand will affect protein rigidity and can lead to physiological responses that are not as desired when the compound is induced. Complexes with an RMSF value < 2 Å are likely to be relatively more stable in the body and can trigger the physiological response of the body (Utari et al., 2025; Shobahah et al., 2025). DiscussionTo the best of our knowledge, this study is the first to explore the potential of natural compounds from the Alpinia genus against the dengue virus, particularly DENV-3. Previous studies have reported that several species within the Alpinia genus possess antiviral potential, such as A. officinarum as a natural protease inhibitor against the Zika virus (Gbore et al., 2023), A. zerumbet for reducing infection by ToMV and TMV (Narusaka et al., 2020), and A. galanga for preventing infection by BVDV (Herrmann et al., 2011). To date, investigations into the antiviral potential of plants from the Alpinia genus are still ongoing, as previous findings have highlighted their promise as sources of natural antiviral agents. In this study, the A. purpurata extract was found to have potential as an anti-dengue agent by inhibiting viral replication in vitro (Table 2). The EC50 values obtained from both the extract and the control reflected the decrease in viral replication activity. This reduction in viral replication activity is presumed to be due to the presence of bioactive compounds, such as flavonoids, terpenoids, and natural glycosides, which can inhibit the active site of the dengue virus protease enzyme. The mechanism of action of the A. purpurata extract is believed to involve binding to the active site of the viral enzyme that encodes replication and repair processes when the virus is damaged by host immune cells (Zhang et al., 2016). The activity concentration pattern against this inhibition agrees with the latest research results on other medicinal plant extracts that show similar activity against DENV-2 and DENV-3. Extracts of Vitex negundo, P. alba, and B. monnieri have been reported to reduce virus titers and prevent the formation of infection foci in Vero cell cultures through a mechanism that inhibits the entry of viruses into cells (Alagarasu et al., 2022). Ansori et al. (2021)showed that natural polyphenolic compounds from Pinus merkusii also exhibit antiviral activity with an IC50 below 150 µg/ml against DENV, confirming the potential of tropical plant secondary metabolites as DENV infection inhibitors (Ansori et al., 2021). The decrease in antiviral activity at high concentrations can also be linked to mild cytotoxic effects, whereby the extract affects cellular metabolism in the host. This can be observed from the tests on Andrographis paniculata and Curcuma longa, which also show similar patterns. Increasing the dose above the optimum limit can reduce the selectivity index and antiviral activity (Ichsyani et al., 2017; Ramalingam et al., 2018). A decrease in the CPE curve in plant extracts may reflect a change in the antiviral mechanism of action from the initial inhibition stage to the virus replication suppression stage. Altamish et al. (2022)showed that extracts containing quercetin and artesunic acid are effective only within a certain concentration range because their mechanism depends on temporary binding to viral surface proteins or the NS3/NS5 enzyme (Altamish et al., 2022). The analysis of natural glycosides from A. purpurata against selected receptors using in silico molecular docking experiments (Table 5) and molecular dynamics simulations (Fig. 5) also supports the in vitro findings obtained earlier. Glycosides were used in this study because previous research has revealed their remarkable potential in triggering important bioactivities within cells. Oleandrin, a natural glycoside derived from Nerium oleander, was found to perfectly bind to essential amino acid residues of viral envelope receptors such as HIV, hepatitis B virus type 1 (HTLV-1), and severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) (Newman et al., 2020). Furthermore, Velandia et al. (2025) demonstrated that the flavonoid class conjugated with sugar moieties in the form of glycosides from 14 herbal plant extracts exhibited stronger inhibitory activity against dengue viral proteins than non-glycosylated flavonoid aglycones (Velandia et al., 2025). In addition to their direct antiviral activity, several studies have shown that glycoside compounds can stabilize cell surfaces and reduce the expression of virus-binding receptors, such as DC-SIGN and CD300a, which play important roles in DENV internalization (Warfield et al., 2016). These studies further strengthen the potential of natural glycosides as safe and abundant antiviral agents in medicinal plants.
Fig. 5. Molecular dynamics of quercetin-3-O-rutinoside complexes. DC-SIGN (CD209) is the main receptor on DCs that mediates DENV attachment through the C-type carbohydrate domain. The docking results show that the quercetin-3-O-rutinoside compound forms strong hydrogen bonds with residues Lys93(A), Glu92(A), Glu364(A), Cys360(A), Ala358(A), and Cys229(A). The formation of these strong hydrogen bonds can inhibit the binding of the mannosylated ligand from the DENV envelope to this host receptor. Hsieh et al. (2020)revealed that increased expression of DC-SIGN enhances DENV infection, suggesting that glycoside binding at this site may inhibit the initial step of viral invasion (Hsieh et al., 2020). Furthermore, a recent study by Xue et al. (2021)supports that the blockade of CD209 by glycoside ligands can reduce the affinity of the pathogen for this receptor, making this mechanism relevant for the inhibition of DENV-3 (Xue et al., 2021). CD300a is an immunomodulatory receptor that regulates phagocytosis and inflammation inhibitory signals. Docking shows that quercetin-3-O-rutinoside binds strongly to residues Glu96(A), Ala98(A), Thr100(A), and Glu121(A) in the CD300a extracellular domain, with a binding energy of -7.1 kcal/mol. This type of interaction can suppress the inhibitory signal of CD300a, which can be exploited by viruses to downregulate cellular immune activation. This finding is supported by the hypothesis that flavonoids can modulate the activity of immune receptors, as in the case of the CD81 receptor, which was successfully blocked by natural flavonoid compounds in an in silico study with a similar interaction pattern on key aromatic residues (Dey et al., 2023). IL-1β acts as the main inflammatory cytokine that increases during DENV infection and is responsible for fever, pain, and vascular permeability. Docking results show that quercetin-3-O-rutinoside occupies the allosteric pocket of the IL-1β protein with a binding energy of -7.2 kcal/mol, forming hydrogen bonds with residues Leu178(A), Tyr184(A), Ser121(A), and Val119(A). Hydrophobic interactions at residues Lys208(A), Lys179(A), and Pro118(A) assist in ligand binding to the receptor. These interactions can actively inhibit the release of IL-1β, which is important for suppressing cytokine storms during severe dengue infection. This is in line with the research conducted by Islam et al. (2025) who reported that several natural compounds with glycoside groups can inhibit proinflammatory cytokines by binding to their catalytic domains (Islam et al., 2025). In addition to its direct antiviral role, A. purpurata extract has the potential to strengthen the oxidative defense system. The compounds quercetin-3-O-rutinoside, sitosteryl-3-O-6-palmitoyl-β-D-glucoside, and kaempferol-3-O-glucuronide were found to form bonds on the active side of SOD that were more stable than the quercetin control. Therefore, natural glycoside compounds can strengthen enzyme stability and reduce oxidative stress in DENV infections. This binding activity agrees with the role of flavonoids in maintaining the stability of antioxidant metalloprotein structures such as SOD and catalase (Pourhajibagher and Bahador, 2024). Previous studies have reported that glycosidic flavonoids such as quercetin-3-O-rutinoside have a hepatoprotective effect through increased antioxidant enzyme activity (Lozano-parra et al., 2025). Thus, this SOD-strengthening effect may contribute to cellular protection against virus-induced oxidative stress. According to the results of the ADMET property screening presented in Table 6, quercetin-3-O-rutinoside was found to be non-mutagenic, non-hepatotoxic, non-skin sensitizing, and did not inhibit the cardiac hERG I channel (only hERG II). These findings are supported by the LD₅₀ value of 1.526 mol/kg and LOAEL value of 2.231 log mg/kg/day, indicating a low acute toxicity category (class 5), as shown in Figure 4. This safety profile represents a major advantage compared with some synthetic antiviral candidates that often cause hepatotoxicity or serious cardiovascular side effects. Furthermore, the absence of hepatic toxicity indicates that this compound is safe for long-term hepatic metabolism. This is particularly important because the liver is the main organ for DENV replication, and a hepatically safe compound would minimize the risk of systemic adverse effects. The total clearance value of 0.187 log mL/min/kg indicates a moderate and stable elimination rate, which may prevent toxic accumulation in the kidneys or other tissues. Although its distribution volume is low (log VDss=−1.597), the relatively high fraction of unbound compound ensures that it remains bioactive in the systemic circulation and can reach target organs such as the liver and spleen. From a neurological perspective, some existing antiviral agents are capable of penetrating the central nervous system, which can result in serious neurological side effects. In this study, the candidate compound showed favorable BBB permeability (−2.215 log BB) and CNS permeability (−4.842 log PS) values, indicating that it does not cross the BBB, thereby reducing the risk of neurotoxicity. Nevertheless, this study highlights a limitation of the compound in terms of its low oral bioavailability, with an intestinal absorption value of approximately 28.49% and a Caco-2 permeability of −0.791 log Papp. This indicates that the compound is poorly absorbed in the gastrointestinal tract. The glycoside has a large molecular weight (>600 Da), high polarity, and multiple hydroxyl and sugar (rutinoside) groups, making it highly polar and less capable of penetrating lipid membranes. Consequently, the systemic effectiveness of the compound when administered orally would be limited. In addition, the compound was identified as both a substrate and an inhibitor of P-glycoprotein, indicating that it can be actively effluxed from intestinal cells by transporter mechanisms, thereby reducing the concentration of the compound that reaches the systemic circulation. The molecular dynamics simulation section validates the molecular docking results by evaluating the stability of the ligand and receptor complex. The results of the MD analysis presented in Fig. 5 indicate that all compounds tend to form stable and well-maintained complexes. However, the complex formed between the DC-SIGN (CD-209) protein and the tested compound exhibited RMSF values above 3 Å, indicating that complex is less stable than the others. The high fluctuations observed in the amino acid residues within the ligand-binding region of the DC-SIGN receptor led to a sharp increase in RMSF values. In addition, the flexible conformation of the active binding site and the high polarity of the compound may have hindered the optimal penetration of the tested ligand into the receptor’s hydrophobic pocket. Huq et al. (2024) showed that the phenolic compound catechin from cacao, which also involves molecular dynamics simulation, exhibits stable bonding with the DENV-3 RdRp enzyme, reinforcing the interpretation of phenolic and biflavonoid complexes as potential antidengue agents in this study. A study reported by Huq et al. (2024) shows that the phenolic compound catechin from cacao, which also involves molecular dynamics simulation, exhibits stable bonding with the DENV-3 RdRp enzyme, reinforcing the interpretation of phenolic and biflavonoid complexes in this study as potential antidengue agents. In addition, the presence of hydrogen, hydrophobic, and Van der Walls interactions in this study formed a strong binding pattern on the amino acid residues present in the body’s immune receptors, which means that it can trigger a bioactivity response to prevent dengue from entering the cell surface. ConclusionIn conclusion, the ethanol extract of A. purpurata shows potential antiviral activity against DENV-3 in vitro and in silico. In vitro testing of viral replication activity showed an EC50 value of 105.8107 µg/mL for the extract, compared to 92.6923 µg/mL for the control. The natural glycoside quercetin-3-O-rutinoside was most effective in inhibiting virus binding to DC-SIGN receptors (−7.8 kcal/mol), balancing immune signals through CD300a receptor regulation (−7.1 kcal/mol), suppressing the production of proinflammatory cytokine IL-1β during a cytokine storm (−7.2 kcal/mol), and activating the endogenous antioxidant superoxide dismutase (−7.0 kcal/mol). Validation using molecular dynamics showed that the overall quercetin-3-O-rutinoside complex tended to be stable with an RMSF value < 3 Å, while the DC-SIGN (CD209) complex had an RMSF value > 3 Å. ADMET and toxicity profiling confirmed favorable pharmacokinetic safety with non-hepatotoxic and non-mutagenic properties. These findings highlight quercetin-3-O-rutinoside as a promising, safe, and multitarget natural antiviral candidate for future in vivo and preclinical development against dengue virus infection. AcknowledgmentsThe authors gratefully acknowledge financial support from the scholarship of Program Magister Menuju Doktor untuk Sarjana Unggul (PMDSU). FundingThis study was supported by the Program Magister Menuju Doktor untuk Sarjana Unggul (PMDSU) scholarship with Decree Number 0419/C3/DT.05.00/2025 and Agreement/Contract Number 059/C3/DT.05.00/PL/2025 from the Ministry of Higher Education, Research, and Technology, Republic of Indonesia. Authors' contributionsConception, design, and materials: MAH, WD, and DW; supervision and resources: WD, DW, and THS; data collection and literature search: MAH, RAR, RJK, and AUR; analysis and interpretation: MAH, AUR, PAW, and FK; manuscript writing: MAH, WD, AUR, RJK, and ER; and critical review: WD, DW, and THS. Conflict of interestThe authors declare no conflict of interest. Data availabilityAll data supporting this study’s findings are available within the manuscript. ReferencesAbdul Ahmad, S.A., Palanisamy, U.D., Tejo, B.A., Chew, M.F., Tham, H.W. and Syed Hassan, S. 2017. Geraniin extracted from the rind of Nephelium lappaceum binds to dengue virus type-2 envelope protein and inhibits early stage of virus replication. Virol. J. 14(1), 229; doi:10.1186/s12985-017-0895-1 Aguinaldo, A., Villaflores, O., Macabeo, A.P., Gehle, D., Krohn, K. and Franzblau, S. 2010. Phytoconstituents from Alpinia purpurata and their in vitro inhibitory activity against Mycobacterium tuberculosis. Pharmacognosy. Mag. 6(24), 339–344. Alagarasu, K., Patil, P., Kaushik, M., Chowdhury, D., Joshi, R.K., Hegde, H.V., Kakade, M.B., Hoti, S.L., Cherian, S. and Parashar, D. 2022. In vitro antiviral activity of potential medicinal plant extracts against dengue and chikungunya viruses. Front. Cellular. InfectMicrobiol. 12, 866452; doi:10.3389/fcimb.2022.866452 Alam, M.M., Jarvis, C.M., Hincapie, R., Mckay, C.S., Schimer, J., Sanhueza, C.A., Xu, K., Diehl, R.C., Finn, M.G. and Kiessling, L.L. 2020. Glycan-modified virus-like particles evoke T helper type 1-like immune responses. ACS. Nano 15(1), 309–321; doi: 10.1021/acsnano.0c03023 Alifiansyah, M.R.T., Herdiansyah, M.A., Pratiwi, R.C., Pramesti, R.P., Hafsyah, N.W., Rania, A.P., Putra, J.E.R.P., Cahyono, P.A., Litazkiyyah, S.K., Muhammad, S.K., Murtadlo, A.A.A., Kharisma, V.D., Ansori, A.N.M., Jakhmola, V., Ashok, P.K., Kalra, J.M., Purnobasuki, H. and Pratiwi, I.A. 2024. QSAR of acyl alizarin red biocompound derivatives of Rubia tinctorum roots and its ADMET properties as anti-breast cancer candidates against MMP-9 protein receptor: in silico study. Food. Syst. 7, 312–320; https://doi.org/10.1016/ Altamish, M., Khan, M., Baig, M.S., Pathak, B., Rani, V., Akhtar, J., Khan, A.A., Ahmad, S. and Krishnan, A. 2022. Therapeutic potential of medicinal plants against dengue infection: a mechanistic viewpoint. ACS. Omega. 7(28), 24048–24065; doi:10.1021/acsomega.2c00625 Ansori, A.N.M., Fadhlolly, A., Proboningrat, A., Hayaza, S., Susilo, R.J.K., Naw, S.W., Posa, G.A.V., Yusrizal, Y.F., Sibero, M.T., Sucipto, T.H. and Seogijanto, S. 2021. In vitro antiviral activity of Pinus merkusii (Pinaceae) stem bark and cone against dengue virus type-2 (DENV-2). Res. J. Pharm. Technol. 1, 3705–3708; doi:10.52711/0974-360x.2021.00641 Aryati, A., Wrahatnala, B.J., Yohan, B., Fanny, M., Hakim, F.K.N., Sunari, E.P., Zuroidah, N., Wardhani, P., Santoso, M.S., Husada, D., Rohman, A., Tarmizi, S.N., Sievers, J.T.O. and Sasmono, R.T. 2020. Dengue virus serotype 4 is responsible for the outbreak of dengue in East Java City of Jember, Indonesia. Viruses 12(9), 913; doi:10.3390/v12090913 Banerjee, P., Kemmler, E., Dunkel, M. and Preissner, R. 2024. ProTox 3.0: a webserver for the prediction of toxicity of chemicals. Nucleic. Acids. Res. 52(W1), W513–W520; doi:10.1093/nar/gkae303 Begum, F., Das, S., Mukherjee, D. and Ray, U. 2019. Hijacking the host immune cells by dengue virus: molecular interplay of receptors and dengue virus envelope. Microorganisms 7(9), 323; doi:10.3390/microorganisms7090323 Carnec, X., Meertens, L., Dejarnac, O., Perera-Lecoin, M., Hafirassou, M.L., Kitaura, J., Ramdasi, R., Schwartz, O. and Amara, A. 2015. The phosphatidylserine and phosphatidylethanolamine receptor CD300a binds dengue virus and enhances infection. J. Virol. 90(1), 92–102; doi:10.1128/JVI.01849-15 Dey, D., Biswas, P., Paul, P., Mahmud, S., Ema, T.I., Khan, A.A., Ahmed, S.Z., Hasan, M.M., Saikat, A.S.M., Fatema, B., Bibi, S., Rahman, M.A. and Kim, B. 2023. Natural flavonoids effectively block the CD81 receptor of hepatocytes and inhibit HCV infection: a computational drug development approach. Mol. Diversity 27(3), 1309–1322; doi:10.1007/s11030-022-10491-9 Fatima, S., Gupta, P., Sharma, S., Sharma, A. and Agarwal, S.M. 2020. ADMET profiling of geographically diverse phytochemical using chemoinformatic tools. Future MedChem. 12(1), 69–87; doi:10.4155/fmc-2019-0206 Gbore, D.J., Zakari, S. and Yusuf, L. 2023. In silico studies of bioactive compounds from Alpinia officinarum as inhibitors of Zika virus protease. Unlocked Informat. Med. 38, 101214. Harapan, H., Michie, A., Yohan, B., Shu, P.Y., Mudatsir, M., Sasmono, R.T. and Imrie, A. 2019. Dengue viruses circulating in Indonesia: a systematic review and phylogenetic analysis of data from five decades. Rev. Med. Virol. 29(4), e2037; doi:10.1002/rmv.2037 Herdiansyah, M.A., Rizaldy, R., Alifiansyah, M.R., Fetty, A.J., Anggraini, D., Agustina, N., Alfian, F.R., Setianingsih, P.N., Elfianah, V., Aulia, H.S., Putra, J.E., Ansori, A.N., Kharisma, V.D., Jakhmola, V., Purnobasuki, H., Pratiwi, I.A., Rebezov, M., Shmeleva, S., Bonkalo, T., Kovalchuk, D.F. and Zainul, R. 2024. Molecular interaction analysis of ferulic acid (4-hydroxy-3-methoxycinnamic acid) as main bioactive compound from palm oil waste against MCF-7 receptors: an in silico study. Narra J. 4(2), e775; doi:10.52225/narra.v4i2.775 Herrmann, F., Romero, M.R., Blazquez, A.G., Kaufmann, D., Ashour, M.L., Kahl, S., Marin, J.J.G., Efferth, T. and Wink, M. 2011. Diversity of pharmacological properties in Chinese and European medicinal plants: cytotoxicity, antiviral and antitrypanosomal screening of 82 herbal drugs. Diversity 3(4), 547–580. Hsieh, T.H., Tsai, T.T., Chen, C.L., Shen, T.J., Jhan, M.K., Tseng, P.C. and Lin, C.F. 2020. Senescence in monocytes facilitates dengue virus infection by increasing infectivity. Front. Cellular Infect. Microbiol. 10, 375; doi:10.3389/fcimb.2020.00375 Huq, A.K.M.M., Roney, M., Dubey, A., Nasir, M.H., Tufail, A., Aluwi, M.F.F.M., Ishak, W.M.W., Islam, M.R. and Tajuddin, S.N. 2024. Phenolic compounds of Theobroma cacao L. show potential against dengue RdRp protease enzyme inhibition by in-silico docking, DFT study, MD simulation and MMGBSA calculation. PLoS One, 19(3), e0299238; doi: 10.1371/journal.pone.0299238 Ichsyani, M., Ridhanya, A., Risanti, M., Desti, H., Ceria, R., Putri, D.H., Sudiro, T.M. and Dewi, B.E. Antiviral effects of Curcuma longa L. against dengue virus in vitro and in vivo. In IOP Conference Series: Earth and Environmental Science, Bristol, UK, IOP Publishing Ltd, 2017, vol. 101(1), p 012005; doi: 10.1088/1755-1315/101/1/012005 Imran, M., Abida., Alotaibi, N.M., Thabet, H.K., Alruwaili, J.A., Eltaib, L., Alshehri, A. and Kamal, M. 2025. Investigation of natural compounds as methyltransferase inhibitors against dengue virus: an in silico approach. J. Biomolecular Struct. &. Dyn. 43(11), 5577–5592; doi:10.1080/07391102.2024.2309647 Islam, M.T., Aktaruzzaman, M., Saif, A., Akter, A., Bhat, M.A., Hossain, M.M., Alam, S.M.N., Rayhan, R., Rehman, S., Yaseen, M. and Raihan, M.O. 2025. In silico-based identification of natural inhibitors from traditionally used medicinal plants that can inhibit dengue infection. Mol. Biotechnol. 67, 2382–2398; doi:10.1007/s12033-024-01204-8 Jasso-Miranda, C., Herrera-Camacho, I., Flores-Mendoza, L.K., Dominguez, F., Vallejo-Ruiz, V., Sanchez-Burgos, G.G., Pando-Robles, V., Santos-Lopez, G. and Reyes-Leyva, J. 2019. Antiviral and immunomodulatory effects of polyphenols on macrophages infected with dengue virus serotypes 2 and 3 enhanced or not with antibodies. Infect. Drug Res. 12, 1833–1852; doi:10.2147/IDR.S210890 Klimoszek, D., Jeleń, M., Dołowy, M. and Morak-Młodawska, B. 2024. Study of the lipophilicity and ADMET parameters of new anticancer diquinothiazines with pharmacophore substituents. Pharmaceuticals 17(6), 725; doi:10.3390/ph17060725 Koishi, A.C., Zanello, P.R., Bianco, E.M., Bordignon, J. and Nunes Duarte Dos Santos, C. 2012. Screening of Dengue virus antiviral activity of marine seaweeds by an in situ enzyme-linked immunosorbent assay. PLos One 7(12), e51089; doi:10.1371/journal.pone.0051089 Kong, L.Y., Zhang, W.J. and Luo, J.G. 2016. The genus Alpinia: a review of its phytochemistry and pharmacology. World. J. Traditional. Chin. Med. 2(1), 26–41. Low, Z.X., Ouyong, B.M., Hassandarvish, P., Poh, C.L. and Ramanathan, B. 2021. Antiviral activity of silymarin and baicalein against dengue virus. Scientific. Rep. 11(1), 21221; doi:10.1038/s41598-021-98949-y Lozano-Parra, A., Herrera, V. and Villar, L.A. 2025. Antioxidant response as a candidate prognostic factor for dengue hypotensive and hemorrhagic complications: results from a nested case-control study in Colombia. Trop. Med. Infect. Dis. 10(1), 14; doi:10.3390/tropicalmed10010014 Mvondo, J.G.M., Matondo, A., Mawete, D.T., Bambi, S.M.N., Mbala, B.M. and Lohohola, P.O. 2021. In silico ADME/T properties of quinine derivatives using SwissADME and pkCSM webservers. Int. J. Trop. Dis. &. Health 42(i1130), 49–92; doi:10.9734/ijtdh/2021/v42i1130492 Newman, R.A., Sastry, K.J., Arav-Boger, R., Cai, H., Matos, R. and Harrod, R. 2020. Antiviral effects of Oleandrin. J. Exp. Pharmacol. 12, 503–515; doi: 10.2147/JEP.S273120 Narusaka, M., Yamaji, Y., Uraji, M., Hatanaka, T., Narusaka. and Y. 2020. Inhibitory effects of Alpinia zerumbet extract against plant virus infection in solanaceous plants. Plant Biotechnol. 37(1), 93–97. Panda, K., Alagarasu, K., Patil, P., Agrawal, M., More, A., Kumar, N.V., Mainkar, P.S., Parashar, D. and Cherian, S. 2021. In vitro antiviral activity of α-mangostin against dengue virus serotype-2 (DENV-2). Molecules. (Basel. Switzerland). 26(10), 3016; doi:10.3390/molecules26103016 Pourhajibagher, M. and Bahador, A. 2024. Bioinformatics analysis of photoexcited natural flavonoid glycosides as the inhibitors for oropharyngeal HPV oncoproteins. AMB. Express. 14(1), 29; doi:10.1186/s13568-024-01684-6 Ramalingam, S., Karupannan, S., Padmanaban, P., Vijayan, S., Sheriff, K., Palani, G. and Krishnasamy, K.K. 2018. Anti-dengue activity of Andrographis paniculata extracts and quantification of dengue viral inhibition by SYBR green reverse transcription polymerase chain reaction. Ayu 39(2), 87–91. Rosmalena, R., Elya, B., Dewi, B.E., Fithriyah, F., Desti, H., Angelina, M., Hanafi, M., Lotulung, P.D., Prasasty, V.D. and Seto, D. 2019. The antiviral effect of indonesian medicinal plant extracts against dengue virus in vitro and in silico. Pathogens (Basel. Switzerland). 8(2), 85; doi:10.3390/pathogens8020085 Sari, A.V., Aryati.., Budhy, T.I., Ma’ruf, A., Husada, D., Palupi, P., Sunari, I.G.A.A.E.P. and Indrasari, Y.N. 2025. Correlation between IL-1β and IL-18 levels with the severity of dengue infection. Indonesian. J. Clin. Pathol. Med. Lab. 31(3), 286–291. Shivanika S Deepak., Kumar V Ragunathan., Tiwari A Sumitha. and Devi, B. 2022. Molecular docking, validation, dynamics simulations, and pharmacokinetic prediction of natural compounds against the SARS-CoV-2 main-protease. J. Biomol. Struct. Dyn. 40, 585–611. Shobahah, J., Wahyuningsih, S., Winarni, D., Herdiansyah, M. and Aly, M. 2025. In silico evaluation of vitamin D3 as a dual inhibitor of CDK2 and Bcl-2 in colorectal cancer. Open. Vet. J. 15(8), 3759; doi:10.1016/j.ovj.2025.03759 Singh, S., Almuhanna, Y., Alshahrani, M.Y., Lowman, D.W., Rice, P.J., Gell, C., Ma, Z., Graves, B., Jackson, D., Lee, K., Juarez, R., Koranteng, J., Muntaka, S., Daniel A. Mitchell., Da Silva, A.C., Hussain, F., Yilmaz, G., Mastrotto, F., Irie, Y., Williams, P., Williams, D.L., Cámara, M. and Martinez-Pomares, L. 2021. Carbohydrates from Pseudomonas aeruginosa biofilms interact with immune C-type lectins and interfere with their receptor function. NPJ. Biofilms. Microbiomes. 7(1), 87; doi:10.1038/s41522-021-00257-w Siti Rahayu., Aulia Umi Rohmatika., Ufairanisa Islamatasya., Mochammad Aqilah Herdiansyah., Raden Joko Kuncoroningrat Susilo., Suhailah Hayaza., Djoko Santoso., Odi Yoshitaka Anggarda., Sri Puji Astuti Wahyuningsih. and Win Darmanto. 2025. Potential of Ganoderma applanatum extract as anticancer and immunomodulator in diethylnitrosamine-induced colon cancer. Trop. J. Natural. Product. Res. 9(9), 4310–4320. Soe, A.M., Ngwe Tun, M.M., Nabeshima, T., Myat, T.W., Htun, M.M., Lin, H., Hom, N.S., Inoue, S., Nwe, K.M., Aye, L.P.P., Fukuta, M., Thant, K.Z., Hasebe, F., Morita, K., Shresta, S., Thu, H.M. and Moi, M.L. 2021. Emergence of a novel dengue virus 3 (DENV-3) genotype-i coincident with increased DENV-3 cases in Yangon, Myanmar between 2017 and 2019. Viruses 13(6), 1152; doi:10.3390/v13061152 Sucipto, T.H., Churrotin, S., Setyawati, H., Kotaki, T., Martak, F. and Soegijanto, S. 2017. Antiviral activity of copper(ii)chloride dihydrate againsts dengue virus type-2 in vero cell. Indonesian J. Trop. Infect. Dis. 6(4), 84–87; doi:10.1016/j.ijtd.2017.01.002 Suzery, M., Ningrum, A.N., Nudin, B., Mulyani, N.S. and Cahyono, B. 2019. Determination of quercetin and rutin in red galangal rhizomes (Alpinia purpurata) and white galangal (Alpinia galanga) with high performance liquid chromatography method. In IOP Conference Series: Earth and Environmental Science, Bristol, UK, IOP Publishing Ltd, 2019, vol. 292(1), pp 12–64; doi:10.1016/j.iosc.2018.01.006 Tanaka, J., Gleinich, A.S., Zhang, Q., Whitfield, R., Kempe, K., Haddleton, D.M., Davis, T.P., Perrier, S., Mitchell, D.A. and Wilson, P. 2017. Specific and differential binding of N-acetylgalactosamine glycopolymers to the human macrophage galactose lectin and asialoglycoprotein receptor. Biomacromolecules 18(5), 1624–1633; doi:10.1021/acs.biomac.7b00228 Titir, S.R., Paul, S.K., Ahmed, S., Haque, N., Nasreen, S.A., Hossain, K.S., Ahmad, F.U., Nila, S.S., Khanam, J., Nowsher, N., Al Amin, A.M.M., Khan, A.U., Aung, M.S. and Kobayashi, N. 2021. Nationwide distribution of dengue virus type 3 (DENV-3) genotype i and emergence of DENV-3 genotype iii during the 2019 outbreak in Bangladesh. Trop. Med. Infect. Dis. 6(2), 58; doi:10.3390/tropicalmed6020058 Utama, I.M.S., Lukman, N., Sukmawati, D.D., Alisjahbana, B., Alam, A., Murniati, D., Utama, I.M.G.D.L., Puspitasari, D., Kosasih, H., Laksono, I., Karyana, M., Karyanti, M.R., Hapsari, M.M.D.E.A.H., Meutia, N., Liang, C.J., Wulan, W.N., Lau, C.Y. and Parwati, K.T.M. 2019. Dengue viral infection in Indonesia: epidemiology, diagnostic challenges, and mutations from an observational cohort study. PLos One 13(10), 7785; doi:10.1371/journal.pntd.0007785 Utari, D., Hayati, A., Winarni, D., Permatasari, I., Fadila, H.A., Qatrunada, N., Rosida, R., Afifudin, A.F.M., Pramudya, M., Muchtaromah, B. and Herdiansyah, M.A. 2025. Biochemical, histological and molecular docking into the impact of polystyrene nanoplastics on antioxidant enzymes and testicular health in rats. J. Anim. Health Prod. 13(3), 729–737. Velandia, S.A., Stashenko, E.E., Quintero-Rueda, E., Conde-Ocazionez, S., Sierra, L.J. and Ocazionez, R.E. 2025. In vitro and in silico analyses explore the role of flavonoid classes in the antiviral activity of plant extracts against the dengue virus. Molecules 30(23), 4566. Victório, C.P., Kuster, R.M. and Lage, C.L.S. 2019. Production of rutin and kaempferol-3-o-glucuronide by tissue cultures of Alpinia purpurata (Vieill) K. Schum. Latin. Am. J. Pharm. 28(4), 613–616. Villaflores, O.B., Macabeo, A.P., Gehle, D., Krohn, K., Franzblau, S.G. and Aguinaldo, A.M. 2010. Phytoconstituents from Alpinia purpurata and their in vitro inhibitory activity against Mycobacterium tuberculosis. Pharmacognosy Magazine 6(24), 339–344; doi: 10.4103/0973-1296.71785 Vora, J., Patel, S., Athar, M., Sinha, S., Chhabria, M.T., Jha, P.C. and Shrivastava, N. 2020. Pharmacophore modeling, molecular docking and molecular dynamics simulation for screening and identifying anti-dengue phytocompounds. J. Biomolecular Struct. Dyn. 38(6), 1726–1740; doi:10.1080/07391102.2019.1615002 Warfield, K.L., Plummer, E.M., Sayce, A.C., Alonzi, D.S., Tang, W., Tyrrell, B.E., Hill, M.L., Caputo, A.T., Killingbeck, S.S., Beatty, P.R., Harris, E., Iwaki, R., Kinami, K., Ide, D., Kiappes, J.L., Kato, A., Buck, M.D., King, K., Eddy, W., Khaliq, M., Sampath, A., Treston, A.M., Dwek, R.A., Enterlein, S.G., Miller, J.L., Zitzmann, N., Ramstedt, U. and Shresta, S. 2016. Inhibition of endoplasmic reticulum glucosidases is required for in vitro and in vivo dengue antiviral activity by the iminosugar UV-4. Antiviral. Res. 129, 93–98; doi:10.1016/j.antiviral.2016.03.001 Xue, Y., Li, Q., Park, C.G., Klena, J.D., Anisimov, A.P., Sun, Z., Wei, X. and Chen, T. 2021. Proteus mirabilis targets atherosclerosis plaques in human coronary arteries via DC-SIGN (CD209). Front. Immunol. 11, 579010; doi:10.3389/fimmu.2020.579010 Zhang, Z., Li, Y., Loh, Y.R., Phoo, W.W., Hung, A.W., Kang, C. and Luo, D. 2016. Crystal structure of unlinked NS2B-NS3 protease from Zika virus. Science 354, 1597–1600; doi: 10.1126/science.aai9309 | ||

| How to Cite this Article |
| Pubmed Style Herdiansyah MA, Darmanto W, Winarni D, Rohmatika AU, Putri RARK, Susilo RJK, Sucipto TH, Wiradana PA, Khanifah F, Ernawati E. Decoding antidengue serotype-3 (DENV-3) mechanism of natural glycosides from Alpinia purpurata: Molecular docking and in vitro insights into viral entry, cytokine storm modulation, and endogenous antioxidant defense activation. Open Vet. J.. 2026; 16(5): 2987-3007. doi:10.5455/OVJ.2026.v16.i5.42 Web Style Herdiansyah MA, Darmanto W, Winarni D, Rohmatika AU, Putri RARK, Susilo RJK, Sucipto TH, Wiradana PA, Khanifah F, Ernawati E. Decoding antidengue serotype-3 (DENV-3) mechanism of natural glycosides from Alpinia purpurata: Molecular docking and in vitro insights into viral entry, cytokine storm modulation, and endogenous antioxidant defense activation. https://www.openveterinaryjournal.com/?mno=304967 [Access: June 26, 2026]. doi:10.5455/OVJ.2026.v16.i5.42 AMA (American Medical Association) Style Herdiansyah MA, Darmanto W, Winarni D, Rohmatika AU, Putri RARK, Susilo RJK, Sucipto TH, Wiradana PA, Khanifah F, Ernawati E. Decoding antidengue serotype-3 (DENV-3) mechanism of natural glycosides from Alpinia purpurata: Molecular docking and in vitro insights into viral entry, cytokine storm modulation, and endogenous antioxidant defense activation. Open Vet. J.. 2026; 16(5): 2987-3007. doi:10.5455/OVJ.2026.v16.i5.42 Vancouver/ICMJE Style Herdiansyah MA, Darmanto W, Winarni D, Rohmatika AU, Putri RARK, Susilo RJK, Sucipto TH, Wiradana PA, Khanifah F, Ernawati E. Decoding antidengue serotype-3 (DENV-3) mechanism of natural glycosides from Alpinia purpurata: Molecular docking and in vitro insights into viral entry, cytokine storm modulation, and endogenous antioxidant defense activation. Open Vet. J.. (2026), [cited June 26, 2026]; 16(5): 2987-3007. doi:10.5455/OVJ.2026.v16.i5.42 Harvard Style Herdiansyah, M. A., Darmanto, . W., Winarni, . D., Rohmatika, . A. U., Putri, . R. A. R. K., Susilo, . R. J. K., Sucipto, . T. H., Wiradana, . P. A., Khanifah, . F. & Ernawati, . E. (2026) Decoding antidengue serotype-3 (DENV-3) mechanism of natural glycosides from Alpinia purpurata: Molecular docking and in vitro insights into viral entry, cytokine storm modulation, and endogenous antioxidant defense activation. Open Vet. J., 16 (5), 2987-3007. doi:10.5455/OVJ.2026.v16.i5.42 Turabian Style Herdiansyah, Mochammad Aqilah, Win Darmanto, Dwi Winarni, Aulia Umi Rohmatika, Rr Aulia Rahmawati Kusuma Putri, Raden Joko Kuncoroningrat Susilo, Teguh Hari Sucipto, Putu Angga Wiradana, Farach Khanifah, and Ernawati Ernawati. 2026. Decoding antidengue serotype-3 (DENV-3) mechanism of natural glycosides from Alpinia purpurata: Molecular docking and in vitro insights into viral entry, cytokine storm modulation, and endogenous antioxidant defense activation. Open Veterinary Journal, 16 (5), 2987-3007. doi:10.5455/OVJ.2026.v16.i5.42 Chicago Style Herdiansyah, Mochammad Aqilah, Win Darmanto, Dwi Winarni, Aulia Umi Rohmatika, Rr Aulia Rahmawati Kusuma Putri, Raden Joko Kuncoroningrat Susilo, Teguh Hari Sucipto, Putu Angga Wiradana, Farach Khanifah, and Ernawati Ernawati. "Decoding antidengue serotype-3 (DENV-3) mechanism of natural glycosides from Alpinia purpurata: Molecular docking and in vitro insights into viral entry, cytokine storm modulation, and endogenous antioxidant defense activation." Open Veterinary Journal 16 (2026), 2987-3007. doi:10.5455/OVJ.2026.v16.i5.42 MLA (The Modern Language Association) Style Herdiansyah, Mochammad Aqilah, Win Darmanto, Dwi Winarni, Aulia Umi Rohmatika, Rr Aulia Rahmawati Kusuma Putri, Raden Joko Kuncoroningrat Susilo, Teguh Hari Sucipto, Putu Angga Wiradana, Farach Khanifah, and Ernawati Ernawati. "Decoding antidengue serotype-3 (DENV-3) mechanism of natural glycosides from Alpinia purpurata: Molecular docking and in vitro insights into viral entry, cytokine storm modulation, and endogenous antioxidant defense activation." Open Veterinary Journal 16.5 (2026), 2987-3007. Print. doi:10.5455/OVJ.2026.v16.i5.42 APA (American Psychological Association) Style Herdiansyah, M. A., Darmanto, . W., Winarni, . D., Rohmatika, . A. U., Putri, . R. A. R. K., Susilo, . R. J. K., Sucipto, . T. H., Wiradana, . P. A., Khanifah, . F. & Ernawati, . E. (2026) Decoding antidengue serotype-3 (DENV-3) mechanism of natural glycosides from Alpinia purpurata: Molecular docking and in vitro insights into viral entry, cytokine storm modulation, and endogenous antioxidant defense activation. Open Veterinary Journal, 16 (5), 2987-3007. doi:10.5455/OVJ.2026.v16.i5.42 |

