Open Veterinary Journal, (2026), Vol. 16(5): 3067-3079
Research Article
10.5455/OVJ.2026.v16.i5.49
Molecular characterization of Echinococcus granulosus isolated from cattle using multilocus genetic markers in Al-Diwaniyah City, Iraq
Marwa Saleem Hajeel and Monyer Abdulamier Abd Alfatlawi*
Department of Veterinary Microbiology, College of Veterinary Medicine, University of Al-Qadisiyah, Al-Diwaniyah, Iraq
*Corresponding Author: Monyer Abdulamier Abd Alfatlawi. Department of Veterinary Microbiology, College of Veterinary Medicine, University of Al-Qadisiyah, Al-Diwaniyah, Iraq. Email: monyerr.abd [at] qu.edu.iq
Submitted: 17/11/2025 Revised: 22/02/2026 Accepted: 02/03/2026 Published: 31/05/2026
© 2025 Open Veterinary Journal
This is an Open Access article distributed under the terms of the Creative Commons Attribution-Non Commercial-No Derivatives License (http://creativecommons.org/licenses/by-nc-nd/4.0/), which permits non-commercial re-use, distribution, and reproduction in any medium, provided the original work is properly cited, and is not altered, transformed, or built upon in any way.
Abstract
Background: Cystic echinococcosis is a major zoonotic parasitic disease that affects livestock and public health worldwide.
Aim: This study aimed to molecularly characterize Echinococcus granulosus from cattle using multilocus polymerase chain reaction (PCR) and deoxyribonucleic acid (DNA) sequencing from Al-Diwaniyah City, Iraq, to understand the unique local variants.
Methods: An examination of 150 slaughtered cattle was conducted in an abattoir in Al-Diwaniyah, Iraq, from January to June 2024. Thirty cattle had fertile hydatid cysts. One cyst was collected from each of the 30 cattle. The germinal layers were used to extract the processed DNA. TEN representatives were analyzed through multilocus PCR and sequenced for ACT-2, cytochrome c oxidase subunit 1 (COX1), cytochrome b (CYT), and NADH dehydrogenase subunit 1 (ND1). This was followed by BLAST and phylogenetic analysis.
Results: Of the 150 inspected cattle, 30 (20.0%) weighed livestock had viable hydatid cysts, and one cyst was harvested per livestock. Multilocus PCR had a success rate of 100% (10/10) for four selected markers (ACT-2, COX1, CYT, and ND1) in the respective isolates. When sequenced, the isolates’ nucleotides matched the global references for E. granulosus within the range of 94.96%–99.61%(mean=96.95%) for ACT-2, 94.87%–98.72% (mean=96.41%) for COX1, 99.54%–100% (mean=99.73%) for CYT, and 96.58%–100% (mean=98.93%) for ND1. Based on the individual marker sequences, phylogenetic construction of the Iraqi cattle isolates clustered them in the E. granulosus sensu stricto complex (G1–G3) with trace amounts of polymorphisms, evidencing a potential for regionally endemic low-diversity haplotypes.
Conclusion: This study shows that cattle in Al-Diwaniyah Province, Iraq, have E. granulosus sensu stricto genotypes (G1–G3) that suggest the presence of local variants despite having overall high sequence similarity and some divergence, particularly at the COX1 locus. The Iraqi cattle isolates being in the same cluster as the regional and global reference strains indicates that the isolates share transmission lineages and have not undergone divergent evolution. This study corroborates the epidemiological finding that cattle are intermediate hosts in dog-mediated transmission cycles and offers country-specific molecular evidence to support the One Health surveillance and control strategies in Iraq.
Keywords: Alignment, Blast, Cattle, Sequencing, Tree.
Introduction
Cystic echinococcosis (CE) is a disease of global importance and is a large zoonotic parasitic disease with significant implications for public health and the health of livestock production systems. Cystic Echinococcosis occurs as a result of the Echinococcus granulosus sensu larval stage. This disease occurs in the Middle East, South America, Africa, and some Asian countries. Traditional animal husbandry systems with continuous transmission cycles occur between the livestock and canids in areas where CE is present (Woolsey and Miller, 2021; Wen et al., 2019). Cystic echinococcosis is a disease that occurs in cattle production systems and results in economic losses due to the condemnation of the infected organs at slaughter, which also reduces the value of the entire slaughtered animal and causes other indirect effects. CE is a large and continuous burden to veterinary and agricultural systems (García-Méndez et al., 2022; Manterola et al., 2022).
Echinococcus granulosus has a rather complex life cycle because it is considered a multi-host parasite. It uses both domestic and wild canids as its definitive hosts. It also has a few intermediate hosts. These include herbivorous mammals such as sheep, cattle, camels, and occasionally humans. During the hydatid cyst stage of the life cycle, humans can become infected mainly in the liver and lungs (Ohiolei et al., 2022). Despite long-standing cattle epidemiological notions, more recent studies have shown that cattle play a positive and more active role in parasite transmission. This has been especially true in endemic areas with poor offal disposal, free-roaming dogs, and slaughterhouse waste (Larrieu et al., 2019; Tamarozzi et al., 2020). Therefore, documenting the genetic variation of E. granulosus in cattle is critical for epidemiological studies.
The different species and genotypes belonging to the E. granulosus complex include E. granulosus sensu stricto (s.s.; G1–G3), Echinococcus equinus (G4), and Echinococcus ortleppi (G5), which pertain to different host transmission tendencies (Khademvatan et al. 2019; Casulli et al. 2022). Echinococcus granulosus complex subsets G1 and G5, respectively, embody sheep-dominated and cattle-dominated host cycles; however, recent studies indicate an increasing overlap of host phenotypes, especially within cattle and other livestock species containing ovine-associated genotypes (Shams et al. 2022; Tamarozzi et al. 2020). These studies prompt a reassessment of the phenotypic specification of dominant livestock and underscore the importance of accurate molecular discrimination in endemic areas.
The molecular characterization of E. granulosus has focused on mitochondrial markers, including cytochrome c oxidase subunit 1 (COX1), cytochrome b (CYT), and NADH dehydrogenase subunit 1 (ND1). These markers are highly discriminatory for genotype and phylogeographic analysis (Hua et al., 2022; Borhani et al., 2024). However, mitochondrial loci alone are insufficient for intraspecific resolution or for distinguishing closely related lineages. The combination of nuclear markers, such as actin genes (ACT), with mitochondrial loci has been shown to provide better resolution of genotypes and stronger molecular epidemiological inferences (Gharbi and Giraudoux, 2024; Rojas-Pincheira et al., 2022). Therefore, multilocus analysis is often regarded as the best option for studying the population structure of E. granulosus.
CE is recognized as endemic in Iraq, yet the molecular data concerning livestock E. granulosus remains sparse and fragmented. The majority of available studies have concentrated almost solely on single mitochondrial markers and on hosts other than cattle, which offers an incomplete understanding of parasite diversity and transmission dynamics. Studies of Iraq’s parasitological landscape and neighboring countries are still few and far between, leaving scholars in the field with limited comprehension of the transboundary parasitological landscape, the movement of parasites, and the extent of their gene flow (Casulli et al., 2022; Hua et al., 2022). Within a One Health framework, these blind spots are critical as integrated control measures call for an understanding of the parasitic genotype(s) present in the animal population, along with the implications for people (Larrieu et al., 2019).
Consequently, the aim of this research is to accomplish a multilocus molecular characterization of E. granulosus isolates from cattle in Al-Diwaniyah Province, Iraq, which entails working with two mitochondrial (COX1 and CYT) and two nuclear (ACT-2 and ND1) molecular markers, and to analyze sequence variability and relatedness to global reference strains through the constructed phylogenetic trees, provide regionally relevant data for epidemiological surveillance, and support one health-based control measures for cystic echinococcosis in Iraq.
Materials and Methods
Sample collection
At an abattoir (Al-Diwaniyah City, Iraq; January and June 2024), hydatid cyst samples were collected from the carcasses of cattle [150 slaughtered cattle; 30 animals (20.0%) were infected] that had been inspected postmortem. Only cysts (fertile based on microscopic confirmation of viable protoscolices) with intact laminated and germinal layers were selected. The cysts were aspirated using sterile syringes, and the germinal layer was removed using sterile forceps and scissors by laminar flow to ensure sterile conditions. The removed germinal layers were placed in sterile Eppendorf tubes. The samples were kept cold until they arrived at the laboratory for processing within a few hours. Upon entering the molecular biology laboratory, the samples were stored at −20°C until required. Freezing the samples was required to stabilize them for downstream molecular biology procedures by preventing deoxyribonucleic acid (DNA) degradation. The biosafety of all samples was ensured while samples were processed according to laboratory biosafety regulations. Biosafety procedures were strictly adhered to during molecular analysis and sequencing. Each tube was labeled with the species from which the samples were obtained, the organ that was collected, and the date of collection.
Genomic DNA extraction
Genomic DNA was extracted from cattle-derived hydatid cysts using the QIAamp DNA Mini Kit (Qiagen, Hilden, Germany), following the manufacturer’s protocol. Approximately 200 µl of lysis buffer and 20 µl of proteinase K solution were added to each germinal layer-containing Eppendorf tube. The samples were thoroughly vortexed and incubated at 56°C for 1 hour to ensure complete cell lysis. After incubation, the lysate was centrifuged at 13,000 rpm for 1 minute to pellet the debris, and the clear supernatant was transferred to fresh sterile tubes.
To this lysate, 200 µl of binding buffer was added and mixed, followed by the addition of 200 µl of absolute ethanol. The entire mixture was transferred into a spin column and centrifuged to allow DNA binding to the column membrane. The column was then washed sequentially with 500 µl of washing buffers 1 and 2, followed by a final dry spin to remove ethanol residues. To establish future uses, the remaining purified DNA was stored in its 50 µl elution buffer and then stored in −20°C. For the polymerase chain reaction (PCR) and sequencing, the integrity and purity of the DNA were required.
Fluorometric DNA quantification was performed using Promega’s USA-made devices. For this purpose, we first diluted the 20X stock TE buffer 20 times and formed a 1X TE buffer. Subsequently, we diluted the QuantiFluor® Dye 1:400, prepared a working solution, and stored it for further use. Control and blank were prepared by placing 200 µl of the working solution in 0.5 ml tubes with the dye and adding 2 µl of standard DNA to the control tubes.
Finally, from all cattle samples, 2 µl of DNA extract was pipetted into a PCR tube and added to 200 µl of working dye solution. Then, it was primed according to the manufacturer’s instructions, and the values were obtained. These values indicated whether the DNA was sufficient for PCR to confirm that the purity and concentration of the DNA were acceptable for the process. All cattle samples confirmed the efficacy of the DNA extraction method. As a result, the samples could advance to the target genes’ PCR amplification.
Selection of primers and amplification of
gene segments via PCR
Using Primer-BLAST (NCBI), four primers were manufactured: two for each of the mitochondrial genes COX1 and CYT, and two for the nuclear genes ACT-2 and ND1. Primers have a tolerance of 5°C for melting temperatures ranging from 55°C to 60°C. Primers with an average GC percentage of 40–60 and a target amplification size of 400–800 bp were designed. To limit the amplification of undesired target genes, each primer pair was validated in silico using available cestode sequences.
Standard reaction conditions were applied to the PCR amplifications. Each reaction was accompanied by a negative control (no DNA template was included). A series of genomic DNA dilutions was prepared to evaluate PCR sensitivity, with the lowest concentration expected to be ~5 ng DNA per reaction.
In this study, we conducted multilocus analysis on four selected gene targets: ACT-2, COX1, CYT, and ND1. We either designed new primer pairs or grounded our approach on previously validated literature. Each PCR reaction was performed in a 20-µl reaction volume containing 10 µl of master mix, 1 µl each of forward and reverse primers, 6 µl of nuclease-free water, and 2 µl of previously extracted cattle DNA. To ensure the consistency of protocols in all genes, PCR amplifications were performed on a T100 Bio-Rad thermal cycler using the same gene cycling conditions.
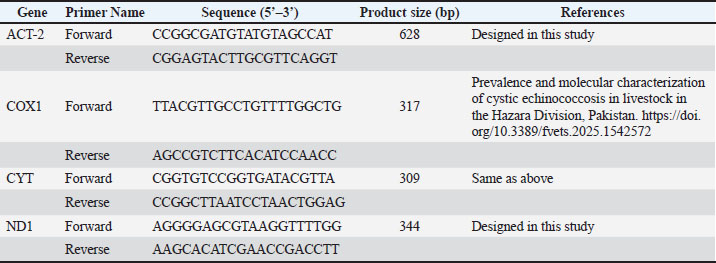
For the thermocycling, the first step was an initial denaturation, which was done for 5 minutes at 95°C, followed by 39 cycles each consisting of a denaturation step done for 30 s at 95 °C, an annealing step done for 35 s at 60 °C, and an extension step done for 35 seconds at 72°C. Extension after 39 cycles was performed at 72°C for 5 minutes. The expected gene segment lengths expected after amplification for the ACT-2, COX1, CYT, and ND1 genes were 628, 317, 309, and 344 bp, respectively. Each PCR was performed alongside a negative control (water in place of the DNA template) to prevent possible contamination during the setup. Table 1 shows the primers used for the different PCRs.
Table 1. Primers used in the current study.
Gel Electrophoresis
The analysis of the PCR products was performed using gel electrophoresis. For each analysis, a gel was prepared at a concentration of 1.5%. This was prepared by mixing 1.5 g of agarose in 100 ml of 1× TBE buffer. The mixture was heated in a microwave until fully dissolved. The mixture was cooled to 60°C, and then 30 l of ethidium bromide was added to the solution. The gel was placed in a casting tray and fitted with a comb. The tray was left at room temperature until fully set, which is approximately 15 minutes. Once set, the comb was removed, and the tray was placed in the electrophoresis tank that was already filled with 1 × TBE buffer.
5 l of each PCR product was then pipetted into the wells, accompanied by a DNA ladder with bands appearing at 100, 150, and 1500 base pairs. Electrophoresis analysis was performed for 1 hour using 100 V. The gel was analyzed using a UV transilluminator connected to a gel doc system that captured the bands for documentation. The PCR products (positive bands) were extracted and purified for sequencing. The products were sequenced to verify that the PCR had amplified the target sequence, in addition to demonstrating that the primers were specific and that amplification had occurred.
DNA sequencing and phylogenetic analysis
The PCR products that had been amplified positively were the samples from the cattle, and 10 of these samples for each gene were selected for sequencing. The Sanger sequencing method was used on the purified amplicons for commercial sequencing, conducted by Macrogen (Korea) and sent through DHL. Chromatograms were returned, and poor-quality signal regions were trimmed. The sequences were cleaned and deposited in GenBank, and the cattle sequences were identified by their corresponding accession numbers.
We ran the cattle sequences through the BLASTn program to identify global matches. Identification of the closest matches was also provided. We will construct the phylogenetic trees with MEGA11, where the maximum likelihood method is applied. The sequences were aligned using ClustalW to determine intra-species variations and local cattle isolates and to determine gaps and substitutions. Finally, the result of the combined platform of analytics provided us with the identification of E. granulosus, which is exemplary of the hinge position, and also the evolution of the cattle in Iraq and the global counterpart.
Ethical approval
Not needed for this study.
Results
ACT-2 Gene
Amplification of the ACT-2 gene from E. granulosus isolated from cattle produced a consistent 628-bp band. All 10 cattle samples showed clear bands of expected size on 1.5% agarose gel. No amplification was observed in the negative control, confirming the specificity of the reaction (Fig. 1).
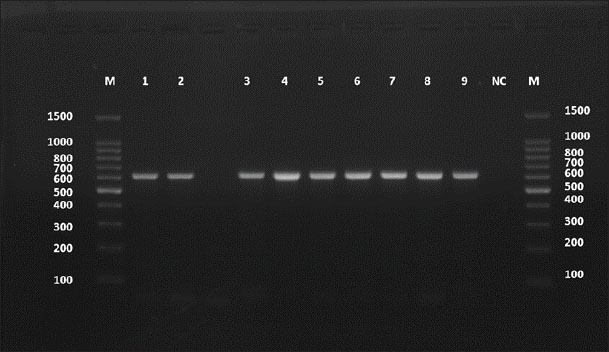
Fig. 1. Agarose gel electrophoresis image (1.5%) shows the amplicons of the ACT2 gene (size=628 bp) in cattle. NC is the negative control in which H₂O was added instead of the template DNA. M is a molecular marker (100–1,500 bp) obtained from GeneDirex (South Korea).
The clear amplification in all samples confirmed that the primers used were effective and that the extracted DNA had good integrity. No nonspecific bands were observed, supporting the specificity of the PCR conditions used.
The ACT-2 gene products from 10 cattle samples were sequenced after PCR. The sequences were submitted to GenBank under the accession numbers PV959514–PV959523. The results of the NCBI-BLAST analyses are presented in Table 2.
Table 2. The NCBI-BLAST Homology Sequence identity (%) between local E. granulosus targeting the ACT2 gene in cattle.
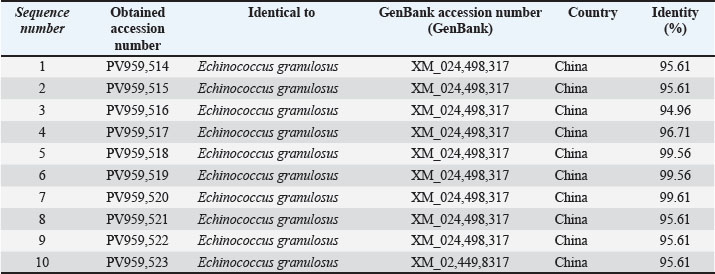
The sequences had high identity with a Chinese reference sequence (XM_024498317) with a range of 94.96%–99.61%, with one sample (PV959520) having the highest identity of 99.61%. Such high similarities attest to the local isolates of E. granulosus sensu stricto (Fig. 2).
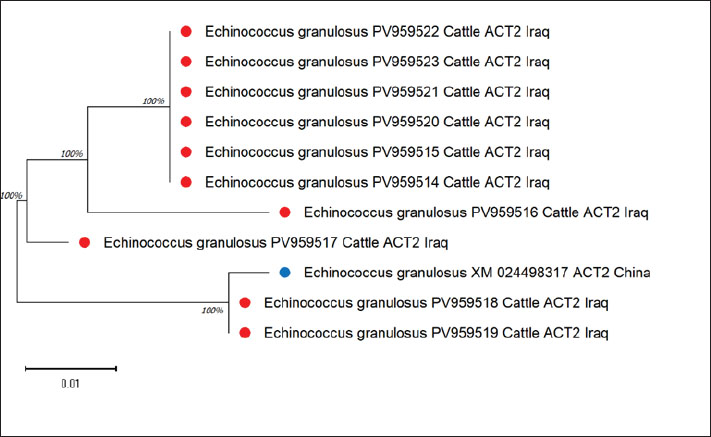
Fig. 2. Evolutionary analysis of Echinococcus granulosus using the maximum likelihood method targeting the ACT2 gene in cattle.
This tree with MEGA11 indicates that the cattle isolates are within the Chinese reference strain, thus supporting the genetic relatedness.
There were only minor differences in single nucleotides, suggesting a low level of genetic variability in the cattle.
COX1 Gene
The PCR amplification of the COX1 gene from cattle-derived isolates registered a sharp, clear 317-bp band across the 10 samples tested, confirming the amplification of the mitochondrial cytochrome c oxidase subunit I gene (Fig. 3).
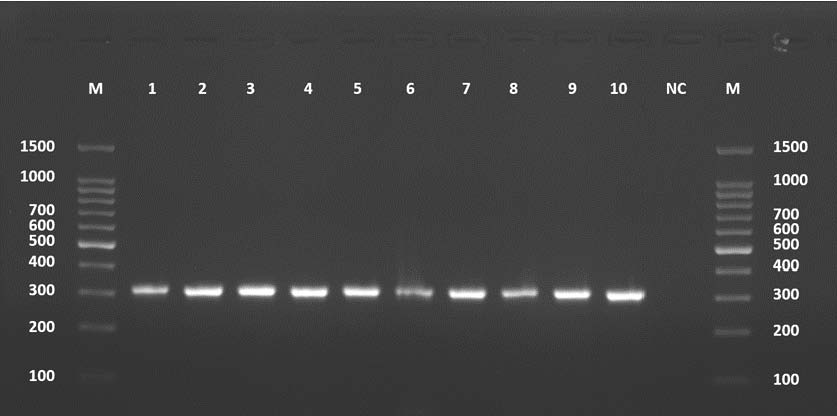
Fig. 3. Agarose gel electrophoresis image (1.5%) showing the COX1 gene amplicons (size=317 bp) in cattle. NC is the negative control in which H₂O was added instead of the template DNA. M is a molecular marker (100–1,500 bp) obtained from GeneDirex (South Korea).
Contamination was unlikely because the negative control had no amplification. The specificity of the primers and optimized PCR conditions were confirmed by yielding expected band sizes across all cattle samples with no extra.
Sequenced PCR products had data that were submitted to GenBank under the accession numbers PV959260–PV959269. The results of BLAST are described in Table 3.
Table 3. The NCBI-BLAST Homology Sequence identity (%) between local E. granulosus targeting the COX1 gene in cattle.
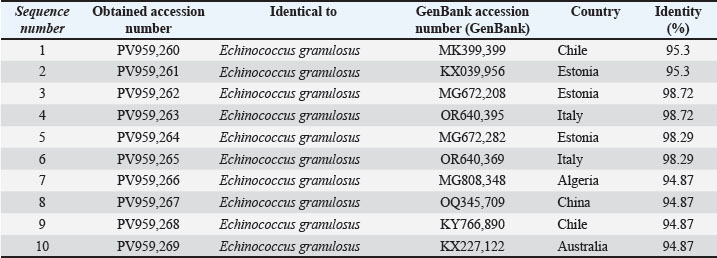
Identity ranged from 98.72% to 94.87%, indicating sequence differences with respect to those from Chile (MK399399), Estonia (MG672208), and Italy (OR640395), which was confirmed by phylogenetic analysis.
Another aspect of this study is that tree analysis showed that the Iraqi cattle isolates were closely related to the European and South American strains (Fig. 4).
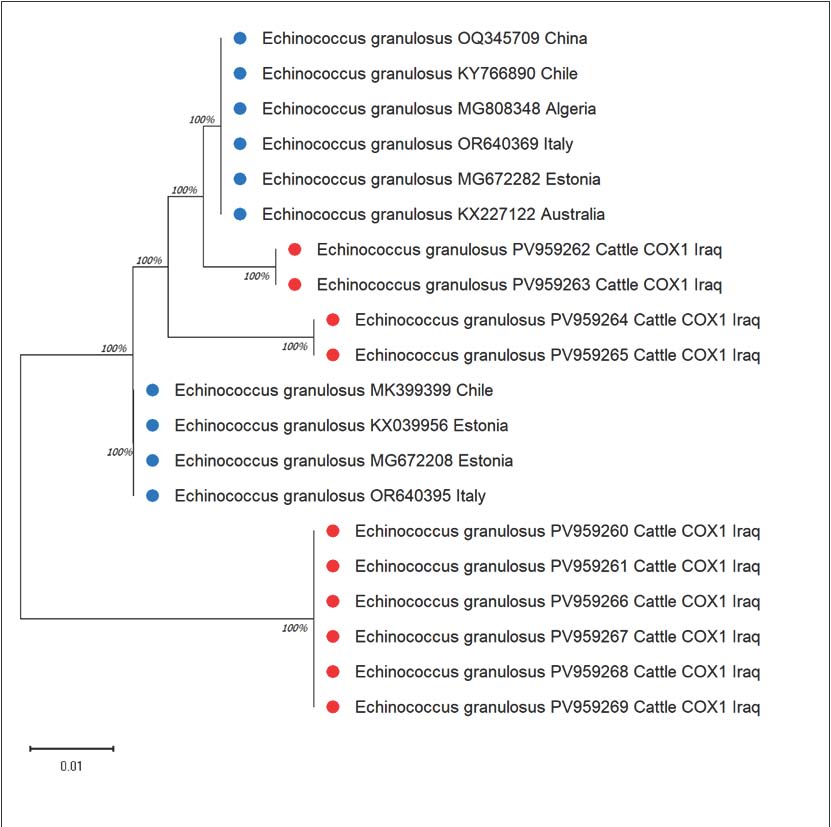
Fig. 4. Evolutionary analysis of Echinococcus granulosus using the maximum likelihood method targeting COX in cattle.
Although the isolates had slight differences in the sequences along the COX1 gene, some degree of separation was observed, suggesting some minor genetic diversity.
CYT Gene
All 10 cattle samples showed CYT gene amplification. This showed a 309-bp band. All PCR results were clean and reproducible.
The amplified bands should be the right size and visible on the agarose gel. No non-specific bands were detected, and no smearing was found, confirming the reliability of the results. The negative results in the control lane show that there was no contamination and that the amplification was not a false positive.
The consistency of the 10 samples shows that the DNA extraction and PCR optimization were performed correctly. The sequences of the cattle CYT gene were deposited in GenBank under the accession numbers PV975181–PV975190. The identity results were found using the BLASTn program, and these results are shown in Table 3.
The percentage of identity of the sequences was 99.54%–100% for reference sequences found in Brazil (MG672224), Pakistan (OP006658), and Estonia (KU925430) (Fig. 5). This shows that cattle isolates are grouped with diverse isolates from around the world, suggesting a low level of divergence (Fig. 6). The presence of polymorphic sites in the local cattle sequences confirms their genetic stability.
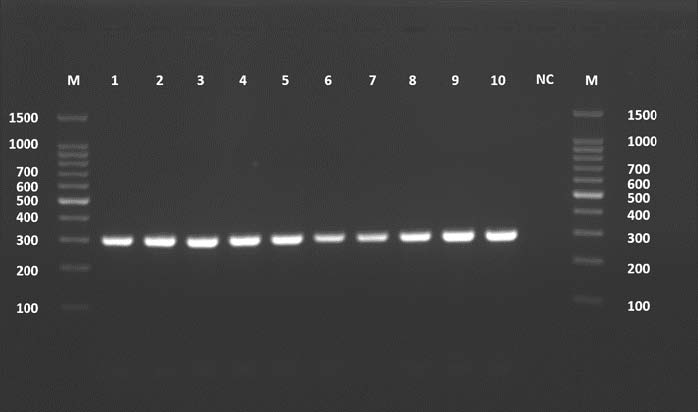
Fig. 5. Agarose gel electrophoresis image (1.5%) shows the amplicons of the CYT gene (size=309 bp) in cattle. NC is the negative control in which H₂O was added instead of the template DNA. M is a molecular marker (100–1,500 bp) obtained from GeneDirex (South Korea).

Fig. 6. Evolutionary analysis of Echinococcus granulosus using the maximum likelihood method targeting the CYT gene in cattle.
ND1 Gene
PCR for the ND1 gene resulted in 344 bp amplicons in all 10 cattle samples. The PCR image showed a strong single band corresponding to the expected product size (Fig. 7).
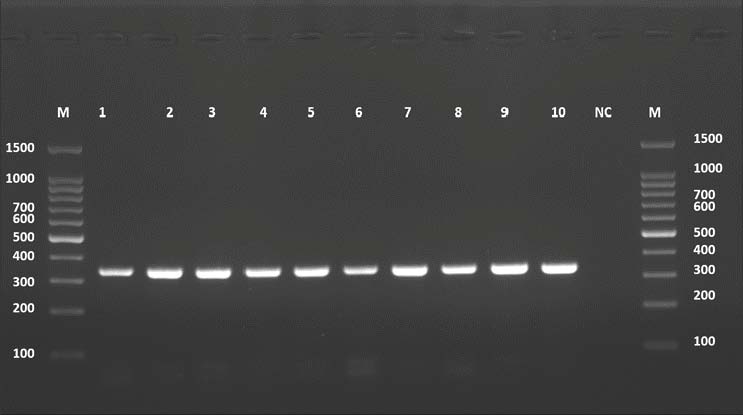
Fig. 7. Agarose gel electrophoresis image (1.5%) shows the COX1 gene amplicons (size=344 bp) in cattle. NC is the negative control in which H₂O was added instead of the template DNA. M is a molecular marker (100–1,500 bp) obtained from GeneDirex (South Korea).
Although the figure label incorrectly refers to COX1, the band size and context confirm that it displays ND1. The presence of clean bands in cattle lanes and the absence of signal in the negative control confirm the specificity of the amplification.
Sequencing of the ND1 gene products resulted in the GenBank entries PV975161–PV975170 (Table 5).
Table 5. The NCBI-BLAST Homology Sequence identity (%) between Echinococcus granulosus targeting the ND1 gene in cattle.
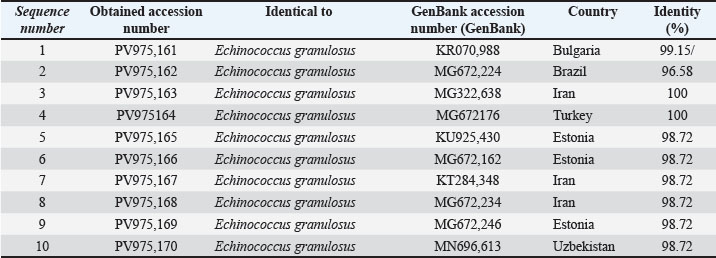
The sequence identity ranged from 96.58% to 100% when compared with isolates from Bulgaria (KR070988), Iran (MG322638), and Turkey (MG672176) (Fig. 8 and Table 4).
Table 4. The NCBI-BLAST Homology Sequence identity (%) between local E. granulosus targeting the CYT gene in cattle.
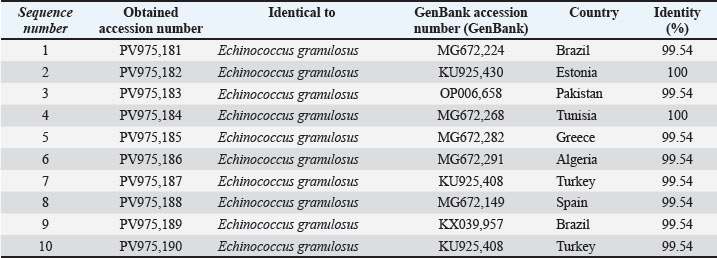

Fig. 8. Evolutionary analysis of Echinococcus granulosus using the maximum likelihood method targeting the ND1 gene in cattle.
Of the 150 cattle examined, 30 (20.0%) were positive for fertile hydatid cysts, from which one cyst per animal was collected, and one cyst from each of the ten animals for multilocus molecular analysis. PCR amplification and sequencing for the four chosen loci (ACT-2, COX1, CYT, and ND1) were successful for all 10 (10/10) of the selected samples. Compared with the GenBank reference strains, ACT-2 (mean 96.95%) presented with 94.96%–99.61% sequence identity, COX1 (mean 96.41%) with 94.87%–98.72%, CYT (mean 99.73%) with 99.54%–100%, and ND1 (mean 98.93%) with 96.58%–100% sequence identity. Low nucleotide polymorphism, few polymorphic sites across loci, and low nucleotide variability suggest a lack of genetic diversity with relative conservation/genetic stability. The phylogenetic reconstruction for all Iraqi cattle isolates within E. granulosus sensu stricto (G1–G3) for both mitochondrial and nuclear markers was consistent using representative reference sequences for established genotypes, supporting the multilocus concordant genotype assignment for the framework. The COX1 nucleotide identity was approximately 95%; however, most of the sequences showed high similarity to global references, suggesting the presence of local variants/sub‐lineages and the absence of technical artifacts. Cattle in the Al-Diwaniyah Province carry ovine-associated E. granulosus genotypes, providing them with epidemiological relevance in local transmission cycles. This evidence is derived from the overall mitochondrial‐nuclear dataset analysis.
Discussion
The cattle isolates from Al-Diwaniyah Province in the present multilocus analysis showed that all of them consistently clustered into the E. granulosus sensu stricto genotypes (G1–G3) on both mitochondrial and nuclear markers, suggesting the existence of a single dominant transmission pattern without the possibility of mixed or host-specific lineages. Although most loci showed high sequence identity, the COX1 identity values that are close to 95% reveal genuine sequence divergence, which is usually attributed to technical error in most studies. However, this explanation is not applicable here because the multiple sequenced loci/genes all aligned well in their respective chromatograms. Cattle involvement in the transmission of the ovine-related Echinococcus genotypes suggests that cattle are not incidental or dead-end hosts in the transmission cycles in this study, which is of considerable importance to livestock epidemiology. It illustrates that there are patterns of shared infection and sustained transmission of infection via dogs in the local livestock systems, a phenomenon indicative of the absence of effective control measures (Sabeeh et al., 2018). From a control perspective, this study suggests that dogs’ access to offal and slaughterhouse practices are potential target to control the transmission of CEC. The study would have benefited from more sequenced isolates and samples from the definitive hosts and infected humans, but nevertheless, the signal of the multilocus sequence data provides a solid baseline for the assessment of the transmission of CEC in cattle in central Iraq.
Findings from the molecular analysis conducted in this study indicated that E. granulosus isolates from infection in cattle in Iraq had significant sequence identity with some E. granulosus strains from China, Brazil, and Iran. This was also documented in some previous genetic reviews, such as that by Hua et al. (2022), where extensive change of genetic variations were documented in E. granulosus in China, predominantly with livestock. In this study, the ACT-2 and ND1 gene segments closely corresponded to accessions from China and Iran, lending credence to the idea of a G1–G3 lineage that is predominantly genetically stable and widely disseminated in farm animals. The proteome of this parasite was also noted by García-Méndez et al. (2022) to have paramount importance globally, which agrees with the findings of high sequence identities and conserved alignments. Although the study was conducted mainly on cattle hosts, some correlating results have also been documented with camels documented that the host species and environmentalvariations determine the hydatid cyst genotypes. This supports the idea that genetic stability in Iraq was a consequence of species overlap with the endemic zone.
Hamad et al. (2025) sequenced E. granulosus antigen B in sheep from Al-Diwaniyah and documented the results closely correlated with the phylogenetic results obtained in cattle using multilocus PCR markers. This could indicate that the strain is conserved in the region as a result of the same circulating dog population and its associated transmission routes.
As Larrieu et al. (2019) noted, livestock-dog cycles dominate CE transmission in rural areas, increasing the likelihood of strain overlap. Koziol and Brehm (2015) also noted the highly conserved nature of E. granulosus genomes, with little variation across the globe, which is consistent with the findings of low divergence in the CYT and COX1 genes. Our phylogenetic analysis corroborated these findings, with the Iraqi cattle isolates clearly showing tight clustering with strains from all over the world. Furthermore, Casulli et al. (2022) noted that the E. granulosus genotypes, particularly with the mixed host populations, showed geographic overlap with no borders. Therefore, it is highly probable that the cattle in Iraq aid in upholding the same strains of the parasite that are also present in sheep and perhaps camels, due to the agricultural characteristics of the region.
The uniformity of the findings from the PCR and sequencing analysis of the samples, especially in the ACT-2, COX1, and ND1 samples, which were identical or nearly identical, showed low inter-regional variation. This is in line with other studies published on Iraq and other regions of the globe, which indicate that the G1-G3 strains are the most prevalent in both cattle and sheep (Koziol and Brehm, 2015; Karim et al., 2018; Abd-Alhassen et al., 2021; Ghazi et al., 2024). Together, these studies strengthen the need for interdisciplinary parasite monitoring and control initiatives in the region.
Conclusion
This study shows that cattle in Al-Diwaniyah Province, Iraq, have E. granulosus sensu stricto genotypes (G1–G3) that suggest the presence of local variants despite having overall high sequence similarity and some divergence, particularly at the COX1 locus. The presence of Iraqi cattle isolates in the same cluster as the regional and global reference strains indicates that the isolates share transmission lineages and have not undergone divergent evolution. This study corroborates the epidemiological finding that cattle are intermediate hosts in dog-mediated transmission cycles and offers country-specific molecular evidence to support the One Health surveillance and control strategies in Iraq.
Acknowledgment
The authors would like to express their sincere appreciation to the College of Veterinary Medicine, University of Al-Qadisiyah, for providing laboratory support and guidance throughout this study.
Conflict of interest
The authors have no conflicts of interest to declare.
Funding
The authors have self-funded the study. No external funding source is available.
Authors’ contributions
All authors have participated in the study.
Data availability
Data are available when requested by the corresponding author.
References
Abd-Alhassen, J.K., Janabi, A.H.D. and Aboktifa, M.A. 2021. Antioxidant and antimicrobial evaluation of lycopene isolated from watermelon. Biochem. Cell. Arch. 21, 2905–2910.
Borhani, M., Fathi, S., Harandi, M.F., Casulli, A., Ding, J., Liu, M., Zhang, W. and Wen, H. 2024. Echinococcus granulosus sensu lato control measures: a specific focus on vaccines for both definitive and intermediate hosts. Parasit. Vectors. 17(1), 533.
Casulli, A., Massolo, A., Saarma, U., Umhang, G., Santolamazza, F. and Santoro, A. 2022. Species and genotypes belonging to Echinococcus granulosus sensu lato complex causing human cystic echinococcosis in Europe (2000–2021): a systematic review. Parasit. Vectors 15(1), 109.
García-Méndez, N., Manterola, C., Totomoch-Serra, A., Riffo-Campos, A.L. and Brito-Carreón, C.A. 2022. Proteomic profile of Echinococcus granulosus: a systematic review. J. Parasitol. 108(1), 64–69.
Gharbi, M. and Giraudoux, P. 2024. Cystic echinococcosis (Echinococcus granulosus sensu lato infection) in Tunisia: a One Health perspective for a future control programme. Parasite 31, 30.
Ghazi, A.M., Ali Al-bayati, M.A. and Janabi, A.H. 2024. Metabolomics-detected alterations generated by phytosomal propolis and phytosomal lycopene in male rats with induced benign prostatic hyperplasia. Iraqi J. Vet. Sci. 38(Suppl. I–IV), 7–15.
Hamad, W.A., Jawad, T.I. and Atiya, W.H. 2025. Phylogenetic analysis and genetic evaluation of the Echinococcus granulosus antigen B subunit 2 gene isolated from sheep in Al-Diwaniyah, Iraq. Open Vet. J. 15(2), 1043–1049.
Hua, R.Q., Du, X.D., He, X., Gu, X.B., Xie, Y., He, R., Xu, J., Peng, X.R. and Yang, G.Y. 2022. Genetic diversity of Echinococcus granulosus sensu lato in China: epidemiological studies and systematic review. Transbound. Emerg. Dis. 69(5), e1382–e1392.
Karim, S., Mansour, K., Janabi, A. and Al-Nakeeb, N. 2018. First phylogenetic characterization of pseudocowpox virus from cattle in Al-Qadisiyah province, Iraq. Iraqi J. Vet. Sci. 33(1), 123–126.
Khademvatan, S., Majidiani, H., Foroutan, M., Hazrati Tappeh, K., Aryamand, S. and Khalkhali, H.R. 2019. Echinococcus granulosus genotypes in Iran: a systematic review. J. Helminthol. 93(2), 131–138.
Koziol, U. and Brehm, K. 2015. Recent advances in Echinococcus genomics and stem cell research. Vet. Parasitol. 213(3–4), 92–102.
Larrieu, E., Gavidia, C.M. and Lightowlers, M.W. 2019. Control of cystic echinococcosis: background and prospects. Zoonoses. Public. Health. 66(8), 889–899.
Manterola, C., Totomoch-Serra, A., Rojas, C., Riffo-Campos, A.L. and García-Méndez, N. 2022. Echinococcus granulosus sensu lato genotypes in different hosts worldwide: a systematic review. Acta Parasitol. 67(1), 161–185.
Ohiolei, J.A., Yan, H.B., Odeniran, P.O., Li, L., Shumuye, N.A., Qurishi, S.A., Isaac, C., Fu, B.Q. and Jia, W.Z. 2022. Echinococcus granulosus sensu lato in animal intermediate hosts: what is with the organ location?. Vet. Parasitol. 304, 109695.
Rojas-Pincheira, C., Manterola, C., García-Méndez, N. and Totomoch-Serra, A. 2022. Spatial distribution of Echinococcus granulosus sensu lato genotypes in the American continent. Rev. Med. Chil. 150(7), 944–957.
Sabeeh, S.A., Esraa, T.M., Alelaah, F.A., Al-Baghdadi, R.J.T. and Janabi, A.H.D. 2018. Metronidazole as a feed additive targeting archaea to reduce frothy bloat in Iraqi cows. Biochem. Cell. Arch. 18(2), 23–28.
Shams, M., Khazaei, S., Naserifar, R., Shariatzadeh, S.A., Anvari, D., Montazeri, F., Pirestani, M. and Majidiani, H. 2022. Global distribution of Echinococcus granulosus genotypes in domestic and wild canids: a systematic review and meta-analysis. Parasitology 149(9), 1147–1159.
Tamarozzi, F., Deplazes, P. and Casulli, A. 2020. Reinventing the wheel of Echinococcus granulosus sensu lato transmission to humans. Trends Parasitol. 36(5), 427–434.
Tamarozzi, F., Legnardi, M., Fittipaldo, A., Drigo, M. and Cassini, R. 2020. Epidemiological distribution of Echinococcus granulosus infection in human and domestic animal hosts in European Mediterranean and Balkan countries: a systematic review. PLos Negl. Trop. Dis. 14(8), e0008519.
Wen, H., Vuitton, L., Tuxun, T., Li, J., Vuitton, D.A., Zhang, W. and Mcmanus, D.P. 2019. Echinococcosis: advances in the 21st century. Clin. Microbiol. Rev. 32(2), e00075–e00018.
Woolsey, I.D. and Miller, A.L. 2021. Echinococcus granulosus sensu lato and Echinococcus multilocularis: a review. Res. Vet. Sci. 135, 517–522.
Yaseen, M.M., Karawan, A.C., Alfatlawi, M.A.A. and Janabi, A.H.D. 2020. Role of gut bacterial cytochrome-P450 of mosquito larvae in degradation of temephos insecticide. Ann. Trop. Med. Public Health 23(1), S412.






