




| Research Article | ||
Open Vet. J.. 2026; 16(5): 2883-2891 Open Veterinary Journal, (2026), Vol. 16(5): 2883-2891 Research Article Comparative effects of raw and commercialized edible bird’s nest products on low-density lipoprotein receptor expression and cholesterol regulation in HepG2 cellsSharifah Zakiah Syed Sulaiman1,2, Mohd Akmal Mohd Noor1, Amirul Nazhan Ilias1, Nurina Titisari3, Ahmad Fauzi1,4 and Mohd Mokrish Mohd Ajat1,5*1Department of Veterinary Preclinical Sciences, Faculty of Veterinary Medicine, Universiti Putra Malaysia, Serdang, Malaysia 2Malaysia Genome and Vaccine Institute, National Institutes of Biotechnology Malaysia, Jalan Bangi, Kajang, Malaysia 3Department of Veterinary Physiology, Faculty of Veterinary Medicine, Universitas Brawijaya, Malang, Indonesia 4Department of Veterinary Clinical Pathology, Faculty of Veterinary Medicine, Universitas Brawijaya, Malang, Indonesia 5Natural Medicine and Product Research Laboratory, Institute of Bioscience, Universiti Putra Malaysia, Serdang, Malaysia *Corresponding Author: Mohd Mokrish Mohd Ajat. Department of Veterinary Preclinical Sciences, Faculty of Veterinary Medicine, Universiti Putra Malaysia, Serdang, Malaysia. Email: mokrish [at] upm.edu.my Submitted: 17/11/2025 Revised: 11/04/2026 Accepted: 23/04/2026 Published: 31/05/2026 © 2025 Open Veterinary Journal
AbstractBackground: Edible bird’s nest (EBN), produced by swiftlets from salivary gland secretions, is a valuable glycoprotein source that offers significant nutritional benefits, particularly influencing consumer health lipid metabolism. Cleaning, bleaching, drying, and enzyme-based techniques can alter the physical quality, nutritional value, and bioactive characteristics of EBN. Aim: This study aimed to compare the effects of raw and commercialized EBN extracts on cell viability, total cellular cholesterol (TCC), and LDLR gene expression in HepG2 cells. Methods: HepG2 cells were cultured under controlled laboratory conditions and treated with graded concentrations of EBN extracts. Cell viability was evaluated using MTT assay. Enzymatically, intracellular cholesterol levels were quantified, and LDLR gene expression was measured using reverse transcription quantitative PCR. Results: Raw EBN maintained higher cell viability and exhibited a nonlinear dose–response pattern, whereas commercialized EBN produced a clear dose-dependent cytotoxic effect. Both raw and commercialized EBN markedly upregulated LDLR gene expression, increasing transcription by approximately 23-fold and 24.5-fold, respectively, compared with untreated controls. Total cellular cholesterol did not differ significantly (p > 0.05), with TCC values of 1.85 ± 0.65, 1.80 ± 0.91, 2.07 ± 0.29, 1.90 ± 0.92, and 3.04 ± 1.04 µmol cholesterol/µmol protein in the BC, NC, PC, raw EBN, and commercialized EBN groups, respectively, although commercialized EBN showed the highest level, representing approximately a 1.6-fold increase relative to BC. Conclusion: Both raw and commercialized EBN significantly enhance LDLR gene expression in HepG2 cells, indicating their potential to promote cholesterol uptake. However, processing may alter the biochemical composition and biological activity of EBN, contributing to differences in cytotoxicity and cholesterol response. These findings provide molecular insights into the potential of EBN as a functional food for cholesterol regulation and highlight the importance of processing integrity in preserving its bioactive efficacy. Keywords: Cholesterol metabolism, Edible Bird’s Nest, HepG2 cells, LDLR gene expression, Lipid regulation. IntroductionEdible bird’s nest (EBN), derived from swiftlet saliva and mainly produced in Southeast Asia, is traditionally valued in Chinese culture as a premium food and health tonic, particularly from A. fuciphagus and A. maximus (Chok et al., 2021). Historically used to enhance vitality, support health, and delay aging, EBN has recently gained scientific attention due to its anti-inflammatory, antioxidant, and bone-supporting effects (Lee et al., 2021). As production shifted from wild cave harvesting to managed swiftlet houses, standardized processing methods such as cleaning, soaking, drying, and packaging became necessary (Ramli et al., 2024). EBN has attracted increasing scientific interest because of its reputation as a traditional health food with various therapeutic benefits (Lee et al., 2023; Acharya and Satheesh, 2023). Biochemically, it is rich in glycoproteins, mainly protein, and contains notable levels of sialic acids, such as N-acetylneuraminic acid, which are associated with its functional effects on brain development, immune regulation, and overall health (Nasir et al., 2021; Ling et al., 2022). Its potential role in lipid metabolism has also attracted attention, with reports demonstrating possible improvements in lipid profiles, adipokine regulation, and insulin sensitivity (Rusanuar et al., 2025). However, despite these promising findings, evidence from large-scale human trials is still lacking, and standardized recommendations regarding dosage and processing have not yet been established. Regulation of cholesterol metabolism can reduce the risk of cardiovascular disease. In humans, the metabolism of cholesterol is predominantly maintained by the liver, where the low-density lipoprotein receptor (LDLR) serves as a major determinant of plasma LDL clearance (Cui et al., 2025). Approximately 70% of circulating LDL is removed via LDLR on hepatocytes, which binds ApoB-containing LDL particles and facilitates their uptake into liver cells (Duan et al., 2022; Li and Li, 2024). Therefore, LDLR expression modulation is a key strategy in managing hypercholesterolemia. Statins lower cholesterol by inhibiting 3-hydroxy-3-methylglutaryl-coenzyme A reductase (HMGCR), the rate-limiting enzyme in cholesterol biosynthesis, which decreases intracellular cholesterol, activates sterol regulatory element binding protein-2 (SREBP-2), and subsequently increases low-density lipoprotein reductase (LDLR) expression to enhance low-density lipoprotein clearance (Shizhan et al., 2019). EBN has attracted increasing attention as a natural regulator of lipid metabolism because it has been shown to increase LDLR expression, suppress HMGCR, improve lipid profiles, reduce hepatic and aortic cholesterol accumulation, and attenuate atherosclerotic lesions in experimental models, with effects comparable to or even greater than simvastatin in some studies (Akmal et al., 2018; Akmal et al., 2020; Nasir et al., 2021). These findings support the potential of EBN as a functional food that modulates cholesterol homeostasis at the molecular level. However, the effect of processing on EBN bioactivity remains unclear, as soaking, boiling, drying, and chemical or heat-based cleaning can alter the physicochemical properties of EBN and affect the structural integrity and potency of key bioactive compounds, such as glycoproteins and peptides (Ramli et al., 2024; Kan et al., 2025). Whole-soup EBN may be more effective than purified extract because it better preserves bioactive compounds in the gelatinous and insoluble fractions (Akmal et al., 2024). Accordingly, this study was conducted to evaluate and compare the effects of raw and commercialized EBN on HepG2 cell viability, LDLR expression, and total cellular cholesterol, with particular emphasis on whether industrial processing modifies its molecular activity related to cholesterol metabolism. Materials and MethodsExperimental designThis study aimed to evaluate and compare the biological effects of raw and commercialized EBN on hepatocyte-like cells, focusing on their impact on cell viability, TCC, and LDLR gene expression. The experiment employed an in vitro model using human hepatocellular carcinoma HepG2 cells, a well-established hepatic model for investigating cholesterol metabolism and lipoprotein receptor activity due to their retention of crucial hepatocyte functions, including lipoprotein synthesis and the expression of genes involved in LDLR regulation and lipid homeostasis (Javitt, 1990; Pramfalk et al., 2016). Five treatment groups were established: baseline control (BC), negative control (NC), positive control (PC; simvastatin-treated), raw EBN-treated (rEBN), and commercialized EBN-treated (cEBN). To ensure reproducibility and statistical validity, all assays were performed in triplicate for each experimental condition. Cell line and culture conditionsHepG2 cells were cultured in AEM (Gibco, USA) supplemented with 10% fetal bovine serum (Gibco), 1% penicillin-streptomycin, and 1% amphotericin B. The cells were maintained in T-75 flasks in a humidified incubator at 37°C and 5% CO₂ until reaching 90% confluency. The culture medium was refreshed every 2 days to maintain optimal growth and prevent nutrient depletion. Sub-culturing was performed under aseptic conditions using 0.25% trypsin-EDTA once the cells reached near confluency. Preparation of raw and commercially available EBN extractsRaw EBN was obtained from a certified manufacturer to ensure the authenticity and quality of the samples. Cleaned EBN were dried for 24 hours in an incubator at 30°C for 24 hours (Memmert, Germany). Dried EBN was ground into powder using a heavy-duty tabletop blender. 1g of EBN powder was mixed with 15 ml of PBS and incubated for 5 h in a 70°C water bath for 5 hours. The EBN was centrifuged at 4°C for 10 minutes at 4400 rpm (Eppendorf, Germany). After centrifugation, the soluble part was collected using a needle-attached syringe and filtered through a 22 µm polyethersulfone syringe filter. Commercially available EBN was purchased and ground into powder using a heavy-duty tabletop blender. The EBN powder was kept at 4°C until further use. Precipitation and quantification of proteinsProtein precipitation from EBN extracts was conducted using the trichloroacetic acid (TCA)-acetone method to isolate soluble proteins for quantification. One volume of EBN extract was mixed with two volumes of cold acetone and incubated at 20°C for 2 hours to promote protein precipitation. The mixture was then centrifuged at 4°C for 30 minutes at 4400 rpm. After discarding the supernatant, the pellet was air-dried to remove residual acetone and subsequently resuspended in chilled PBS (2.0 ml). The suspension was vortexed for 5 minutes and incubated at 4°C overnight to ensure full solubilization. The Bicinchoninic Acid (BCA) Protein Assay (Thermo Fisher Scientific, USA) was used to determine the protein concentration following the manufacturer’s instructions, and the resulting values were used to normalize the cholesterol measurement data. Briefly, 200 µl working reagent was added to 25 µl protein standards and the unknown sample in a microplate. Next, the microplate was incubated for 30 minutes at 37°C for 30 minutes. The absorbance at 560 nm was measured after incubation, and a standard curve was generated to determine the protein concentration in the unknown samples. Cell viability assayCells (3 × 104 cells per well) were seeded in a 96-well plate with raw and commercialized EBN at different concentrations (0.2, 0.5, 0.8, 1.0, and 1.5 mg/ml). After 24 hours of incubation, the treatment media was removed, and 100µl of new media was added. After adding 10 µl of a 5-mg/ml 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) solution, the microplate was wrapped in aluminum foil and incubated for 4 h. After incubation, 100 µl of dimethyl sulfoxide (DMSO) was added to each well, and the media was removed. The absorbance at 570 nm was measured after the microplate was placed on an orbital shaker for 20 minutes. The following formula was used to determine the cell viability percentage:
where A represents the absorbance. The results are expressed as mean ± standard deviation. Cell treatmentFor subsequent biochemical and molecular assays, HepG2 cells were seeded at 1 × 10⁶ cells per well in 6-well plates and allowed to adhere for 24 hours before treatment. The cells were divided into five experimental groups: After that, the cells were treated as follows: (1) Base control: AMEM, 1% FBS; (2) Negative control: AMEM, 1% FBS, 250x cholesterol, 2500 µg/ml LDL; (3) Positive control: AMEM, 1% FBS, 1:250 cholesterol, 10 µg/ml LDL, 4.6 µg/ml simvastatin; (4) rEBN: AMEM, 1% FBS, 1:250 cholesterol, 10 µg/ml LDL, 1.5 mg/ml raw EBN; (5) cEBN: AMEM, 1% FBS, 1:250 cholesterol, 10 µg/ml LDL, and 1.5 mg/ml commercialized EBN. Quantification of total cellular cholesterolFollowing treatment, the treatment media were removed using a portable aspiration system, and cells were washed twice with chilled phosphate-buffered saline (PBS)for 2 times. The cells were scraped using a cell scraper and transferred to another tube. Subsequently, cells were homogenized using an ultrasonic dismembrator at 20% amplitude for 2 minutes. Lipid extraction was performed according to the method of Bligh and Dyer (1959). Total cellular cholesterol was measured using the cholesterol liquichlor kit (Human Diagnostic, USA) according to the manufacturer’s protocol. The absorbance was measured at 540 nm. The TCC was calculated as follows:
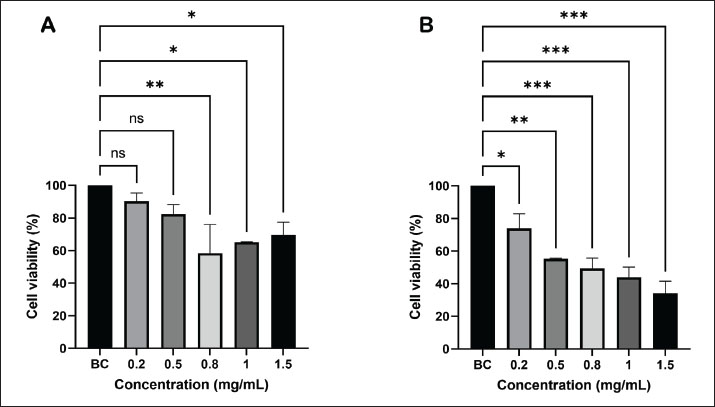
Cholesterol values were normalized to total protein content as determined by the BCA assay to account for cell number variations. LDLR expression analysis using RT-qPCRRibonucleic acid (RNA) extraction was performed using the Qiagen RNeasy® Mini Kit. RNA concentration and purity were measured using the NanoQuant plate and the Infinite M200Pro Spectrophotometer (Tecan, Switzerland). Prior to RT-qPCR, the total RNA content was normalized to 100 ng/µl. RNA amplification was performed using the GoTaq® qPCR Master Mix (Promega, USA). The initial step of gene amplification included the reverse transcription step, which was performed for 15 minutes at 45°C. The reverse transcriptase inactivation and DNA polymerase activation steps were performed for 10 minutes at 95°C. Following that, 30 seconds of denaturation was performed at 94°C, and the annealing stage was performed at 61°C (GAPDH) and 64°C (LDLR) for 1 minutes. Finally, the extension step was performed at 68°C for 2 minutes. The GAPDH and LDLR products were 369 and 453 bp, respectively. The ∆∆CT method was used to quantify target gene expression relative to GAPDH as the internal reference gene. Data are presented as fold changes relative to the baseline control. The primer sequences used for the gene expression study were as follows: for GAPDH, the forward primer was 5’–GAC CAC AGT CCA TGC CAT CAC–3’ and the reverse primer was 5’–TCC ACC ACC CTG TTG CTG TAG–3’, while for LDLR, the forward primer was 5’–CCC CGC AGA TCA ACC CCC ACT C–3’ and the reverse primer was 5’–AGA CCC CCA GGC AAA GGA AGA CGA–3’. Statistical analysisAll quantitative data are expressed as mean ± SD. Statistical analyses were performed using one-way analysis of variance followed by Tukey’s post hoc test to determine pairwise differences between groups. Statistical significance was set at p < 0.05. All experiments were conducted using three independent biological replicates to ensure accuracy and reproducibility. Ethical approvalThis study used a commercially available human cell line (HepG2) without human or animal subjects; therefore, ethical approval was not required. All experiments were conducted following biosafety regulations and good laboratory practice guidelines. Reagents, instruments, and consumables were standardized to minimize variability, and all treatments were freshly prepared before use. Each experimental procedure was performed under sterile conditions to prevent contamination and ensure data reliability. ResultsEffects of different concentrations of raw and commercialized EBN on HepG2 cell viabilityThe cytotoxic profiles of raw and commercialized EBN extracts were analyzed using the MTT assay to identify suitable concentration ranges for further molecular studies, revealing different viability patterns between the two preparations (Fig. 1). The raw EBN showed a nonlinear pattern (Fig. 1A). At 0.2 mg/ml, viability was highest (90.30% ± 5.04%), then decreased slightly at 0.5 mg/ml (82.46% ± 5.61%). A significant decline was observed at 0.8 mg/ml (58.40% ± 17.64%; p < 0.05 compared with the control), indicating moderate cytotoxicity. Interestingly, viability partially recovered at 1.0 mg/ml (65.16% ± 0.15%) and 1.5 mg/ml (69.68% ± 7.74%). Only the 0.8 mg/ml group exhibited a significant difference from the control, indicating that raw EBN was typically well tolerated within this concentration. Commercialized EBN yielded a distinct outcome (Fig. 1B). Here, the viability declined steadily with increasing concentration: 74.07% ± 8.83% (0.2 mg/ml), 55.22% ± 0.42% (0.5 mg/ml), 49.43% ± 6.35% (0.8 mg/ml), 43.88% ± 6.27% (1.0 mg/ml), and 34.25% ± 7.30% (1.5 mg/ml). All treated groups exhibited substantial decreases relative to the control (p < 0.05–0.001), demonstrating a distinct dose-dependent cytotoxic impact. These data indicate that while raw EBN preserves cell viability throughout a broad dosage spectrum and exhibits a biphasic response, commercial EBN demonstrates a more uniform cytotoxic effect. However, both extracts maintained over 50% viability at various concentrations, thereby indicating the use of 1.5 mg/ml for subsequent investigations.
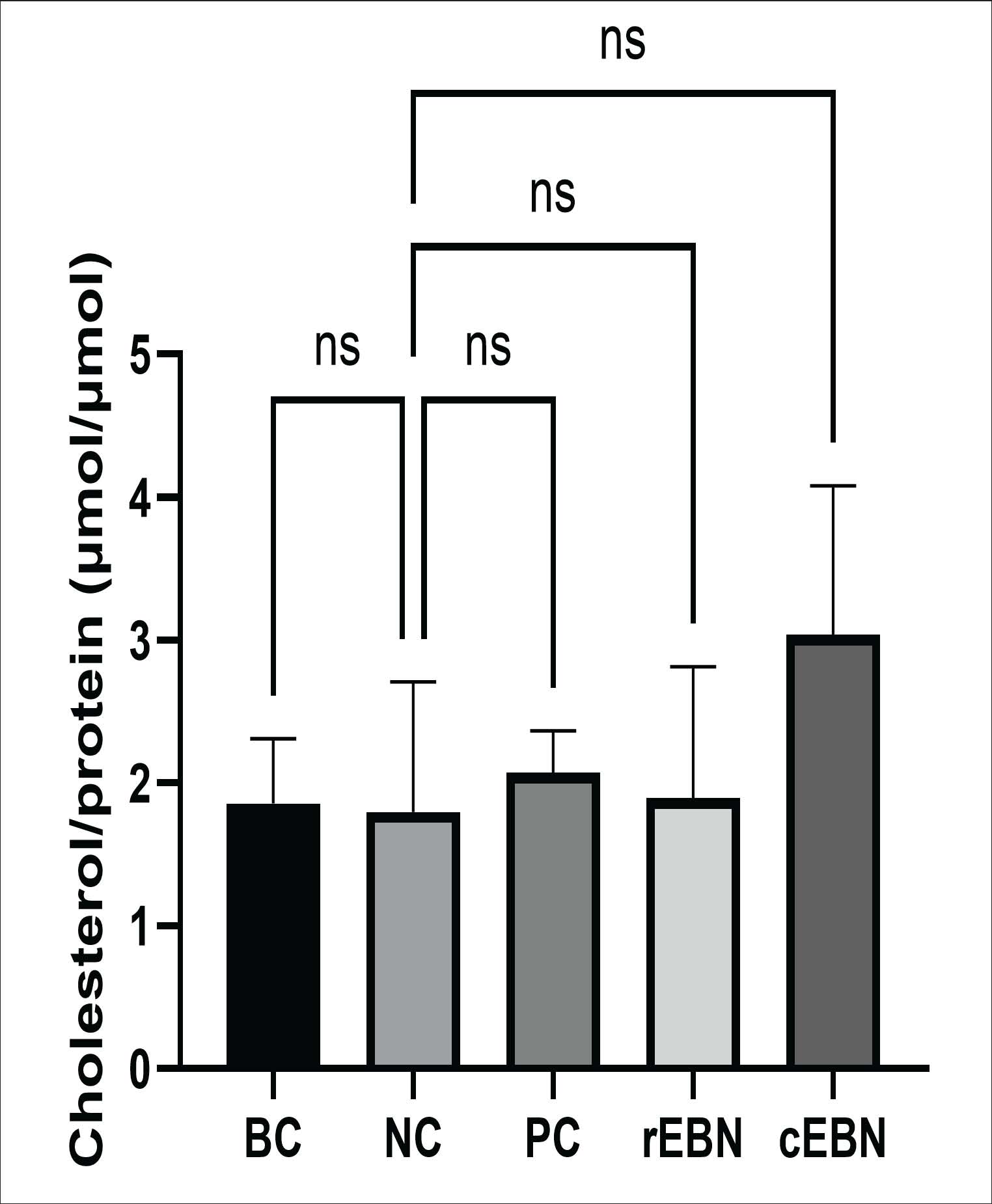
Fig. 1. Viability of HepG2 cells treated with different concentrations of raw (A) and commercialized (B) EBN after 24 hours of treatment. *p < 0.05, **p < 0.01, ***p < 0.000. Effects of raw and commercialized EBN on total cellular cholesterolThe TCC content in HepG2 cells was measured after 24 hours of exposure to each experimental condition to assess the effect of EBN treatments on intracellular lipid homeostasis (Fig. 2). The BC group had a TCC of 1.85 ± 0.65 µmol cholesterol/µmol protein. The negative control showed a similar value (1.80 ± 0.91 µmol cholesterol/µmol protein), indicating that the 24-hours loading protocol did not substantially increase the intracellular cholesterol level in this setting. The positive control yielded a TCC of 2.07 ± 0.29 µmol cholesterol/µmol protein, which was not significantly different from the baseline. Raw EBN (rEBN) produced a TCC of 1.90 ± 0.92 µmol cholesterol/µmol protein, which was very close to that of the control groups. In contrast, cEBN showed the highest TCC (3.04 ± 1.04 µmol cholesterol/µmol protein), equivalent to approximately a 1.6-fold increase relative to BC, although this difference did not reach statistical significance (p > 0.05). Thus, within 24 hours, neither form of EBN significantly changed TCC within 24 hours, but commercialized EBN tended to increase intracellular cholesterol, raising the possibility of altered cholesterol handling in this group.
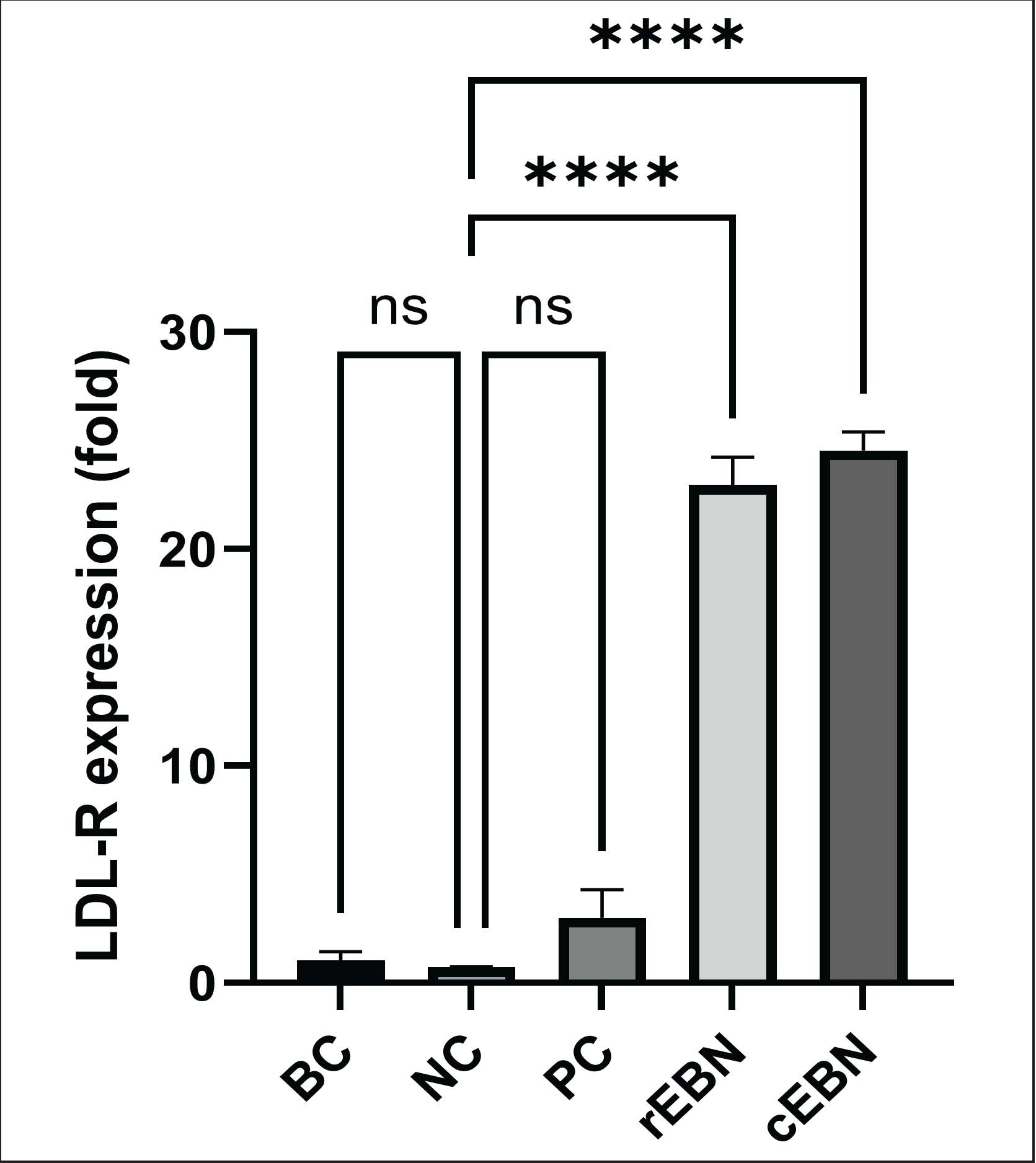
Fig. 2. Total cellular cholesterol (TCC) in the control (BC, baseline control; NC, negative control; PC, positive control) and EBN treatment (rEBN, raw edible bird’s nest; cEBN, commercialized edible bird’s nest) groups. ns: not significant. Effects of raw and commercialized EBN on LDLR gene expressionGene expression data were normalized against the housekeeping gene GAPDH, and fold changes were calculated relative to the baseline control using the ΔΔCt method. Subsequently, LDLR mRNA expression levels were measured to assess the impact of EBN on cholesterol uptake pathways (Fig. 3). The BC and NC groups had low and comparable LDLR expression, consistent with basal levels in untreated and cholesterol-loaded cells. Simvastatin (PC group) produced a modest, non-significant increase, consistent with its mechanism of action. Both the EBN preparations had a much stronger effect. Raw EBN increased LDLR expression by approximately 23.0 ± 2.5-fold (p < 0.001 vs BC group) and commercialized EBN by about 24.5 ± 3.1-fold (p < 0.001 vs BC group). These increases were significantly higher than those in the control and simvastatin groups. The difference between raw and commercialized EBN was not statistically significant, indicating that both forms are similarly potent in upregulating LDLR transcription despite their different effects on viability and TCC.
Fig. 3. Expression of the LDLR gene in HepG2 cells in the control (BC, baseline control; NC, negative control; PC, positive control) and EBN treatment groups (rEBN, raw edible bird’s nest; cEBN, commercialized edible bird’s nest). DiscussionThis study investigated the comparative effects of raw and commercialized EBN on HepG2 cells, focusing on cell viability, TCC, and LDLR gene expression. The findings demonstrate that both forms of EBN markedly upregulated LDLR expression but exhibited distinct effects on cell viability and intracellular cholesterol accumulation. Raw EBN exhibited a mild and nonlinear cytotoxic response with minimal toxicity, whereas commercialized EBN exhibited a clearer concentration-dependent decline in cell viability and a trend toward higher cholesterol content. These results confirm that both raw and processed EBN possess active compounds capable of modulating lipid metabolism. However, processing-related alterations may influence their biochemical activity and cellular responses. The results of cell viability assays highlight the cytocompatibility of the effects of raw and commercialized EBN on HepG2 cells. Raw EBN exhibited a biphasic pattern of cell viability, increasing at low concentrations, decreasing at intermediate levels, and increasing again at higher concentrations, showing that different doses activate distinct cellular pathways. Similar findings by Alexa et al. (2022) on a natural preparation reveal stimulating effects at low doses, significant cytotoxicity at moderate doses, and resurgence in viability at higher doses, particularly noted in the combination of BER and OEO (Alexa et al., 2022). In contrast, commercialized EBN displayed a clear dose-dependent cytotoxic effect, with decreased cell viability correlated with increased EBN concentrations. A growing body of evidence indicates that natural products often exhibit concentration-dependent cytotoxic effects on cultured cells, where the biological response varies significantly across different dose ranges. Bairagee et al. (2025) identified multiple pathways activated by herbal medicines, leading to enhanced cytotoxic outcomes as exposure levels rise (Bairagee et al., 2025). As summarized in the review by Gavanji et al. (2023), many herbal-derived compounds modulate cellular viability through mechanisms such as oxidative stress, apoptosis, mitochondrial dysfunction, and DNA damage, with these effects intensifying as the extract concentration increases (Gavanji et al., 2023). For instance, low concentrations of R. communis leaf extracts showed little to no toxicity in sel RAW 264.7, whereas higher concentrations (≥500 µg/ml) caused a marked decline in cell viability, confirming a direct concentration–cytotoxicity relationship under in vitro conditions (Nemudzivhadi and Masoko, 2014). Variations in cell viability across raw and commercialized EBN may be attributed to the concentrations of bioactive compounds. This may differ depending on the source, species, habitat, and industrial processing steps applied, as suggested by several studies (Lee et al., 2021; Zulkefe et al., 2024). For example, prolonged incubation of EBN in simulated gastric conditions (pepsin, pH 2) promotes the release of Neu5Ac from its conjugated forms, resulting in higher detectable Neu5Ac levels and enhanced bioactivities compared with untreated EBN samples (Chok et al., 2021). The stronger cytotoxicity of commercialized EBN may reflect processing-induced degradation, underscoring the need to preserve biochemical integrity. The TCC results showed that neither EBN form significantly altered intracellular cholesterol content, demonstrating that lipid modulation occurs primarily through transcriptional regulation rather than immediate biochemical changes. The slight, non-significant increase in TCC observed with commercialized EBN may indicate enhanced LDL uptake coupled with less efficient cholesterol use or efflux, whereas raw EBN maintained levels close to baseline. EBN downregulates HMG-CoA reductase, the rate-limiting enzyme in cholesterol biosynthesis, thereby modulating hepatic cholesterol metabolism. The suppression of HMGCR is consistent with the observed improvements in lipid profile, indicating that EBN may reduce endogenous cholesterol synthesis and contribute to more favorable metabolic regulation (Permatasari et al., 2023). In contrast, simvastatin, a cholesterol-lowering medication, primarily inhibits HMGCR, thereby reducing cholesterol synthesis. This leads to decreased total cholesterol levels in HepG2 cells (Verma et al., 2022). The mechanism of simvastatin is aimed at lowering cholesterol production, which is fundamentally different from the action of commercialized EBN. Interestingly, the results of raw EBN were similar to those of simvastatin, with no significant increase in total cholesterol levels compared to the commercialized EBN. This indicates that the bioactive components of raw EBN may regulate cholesterol synthesis, utilization, and degradation, comparable to statins, which inhibit HMGCR and elevate LDLR expression to enhance cholesterol uptake and clearance. The processing methods used to produce commercialized EBN may alter its effects on cholesterol metabolism, leading to the observed increase in total cholesterol levels (Yida et al., 2022). The significant upregulation of LDLR gene expression in HepG2 cells treated with both EBN preparations is a major outcome of this study. By increasing its presence on hepatocyte surfaces, LDLR is the main regulator of plasma LDL-cholesterol clearance, which enhances the binding and internalization of circulating LDL particles, thereby facilitating cholesterol removal from the bloodstream (Aldworth and Hooper, 2024). The results revealed ~23-fold and ~24.5-fold increases in LDLR expression for raw and commercialized EBN, respectively. This suggests that EBN contains molecular constituents that can activate LDLR-related transcriptional pathways, supporting earlier findings by Akmal et al. (2020) on HMGCR suppression and increased LDLR expression (Akmal et al., 2020). The comparable levels of LDLR induction observed for both raw and commercialized EBN show that the bioactive molecules responsible for LDLR upregulation may remain partially preserved during processing. EBN processing plays a decisive role in shaping its biological efficacy. Industrial processing steps, including heating, drying, and other post-harvest treatments, induce physicochemical modifications in EBN (Chok et al., 2021; Unal et al., 2022). These procedures can reduce sialic acid retention, alter amino acid composition, and potentially affect the glycosylation patterns of EBN glycoproteins, thereby modifying their nutritional and biological properties (Gan et al., 2020). Such structural changes may diminish antioxidant capacity or disrupt glycoprotein-cell interactions. The stronger cytotoxicity and higher TCC trend observed in commercialized EBN-treated cells may be due to bioactive profiles modified by processing or reduced molecular stability. In contrast, raw EBN with minimal processing retains its natural biochemical integrity and demonstrates superior cellular compatibility. These findings reinforce the notion that processing is a critical determinant of the nutraceutical potential of EBN, influencing its efficacy and safety. From a practical perspective, standardizing EBN production with controlled temperature, pH, and drying conditions may help preserve its bioactive components and ensure consistency in biological effects. The findings indicate that EBN may serve as a natural modulator of lipid metabolism via the LDLR pathway, while also highlighting the need to assess the effects of processing methods on its safety and efficacy. Raw EBN shows promise for nutraceutical applications due to better cellular tolerance and lipid regulation. Future studies should focus on identifying compounds in EBN that activate LDLR, including glycoproteins and sialic acid derivatives, and how processing influences these compounds. Proteomic and metabolomic analyses could also reveal structural changes that affect functionality. Although HepG2 cells are a recognized model, in vivo studies are necessary to verify whether LDLR upregulation leads to improved systemic lipid profiles and to explore the long-term implications on liver function and cholesterol management by assessing HMGCR, SREBP2, and PCSK9. ConclusionBoth raw and commercialized EBN extracts significantly increased LDLR gene expression in HepG2 cells, indicating their potential to enhance cholesterol metabolism. However, raw EBN maintained higher cell viability and stable cholesterol levels, whereas commercialized EBN exhibited cytotoxicity and increased cellular cholesterol due to processing. These findings highlight the bioactive potential of EBN for lipid regulation and raise concerns regarding the impact of industrial processing. EBN may therefore serve as a potential functional food with cholesterol-regulating properties similar to those of statins. Nevertheless, further investigations, including compositional analysis, protein validation, and in vivo studies, are essential to understand the mechanisms of EBN in lipid metabolism and to develop processing methods that maintain its bioactive qualities. AcknowledgmentThe authors would like to thank the Faculty of Veterinary Medicine, Universiti Putra Malaysia, for providing the facilities to conduct this study. Conflict of interestThe authors declare that there are no conflicts of interest regarding the publication of this manuscript. FundingThis project was financially supported by the UPM RMC IP Grant (GP/2020/9693000). Authors' contributionsSharifah Zakiah Sulaiman performed the experiments, analyzed the data, and drafted the manuscript. Mohd Akmal Mohd Noor and Amirul Nazhan Ilias performed the laboratory work and data acquisition. Nurina Titisari contributed to data interpretation and manuscript editing. Ahmad Fauzi provided methodological support and critical revisions. Mohd Mokrish Mohd Ajat supervised the project, secured funding, and approved the final manuscript. Data availabilityThe data that support the findings of this study are available upon request from the corresponding author. ReferencesAcharya, C. and Satheesh, N. 2023. Edible bird’s nest (EBN): production, processing, food and medicinal importance. AgriCos E-Newslett. 4(3), 20–23. Akmal, M., Razak, I.S.A., Ajat, M., Mansor, R., Zakaria, M.Z.A.B. and Ideris, A. 2018. Edible bird’s nest (EBN) supplementation ameliorates the progression of hepatic changes and atherosclerosis in hypercholesterolaemic-induced rats. Malays. J. Microsci. 14, 103–114. Akmal, M., Razak, I.S.A., Ajat, M., Zakaria, M.Z.A.B., Mansor, R., Ideris, A. and Omar, A.R. 2024. Edible bird’s nest soup (EBNS) serves as an anti-obesity and antilipemic after 6 weeks of supplementation in rats fed a high-fat diet (HFD). Sains Malaysiana 53(11), 3545–3553. Akmal, M.N., Abdul Razak, I.S., Mansor, R., Ideris, A., Omar, A.R., Abu, J. and Ajat, M. 2020. High-dose edible bird’s nest extract (EBN) upregulates LDL-R via suppression of HMGCR gene expression in HepG2 cell lines. Sains. Malaysiana. 49(10), 2433–2442. Aldworth, H. and Hooper, N.M. 2024. Post-translational regulation of low-density lipoprotein receptors provides new targets for cholesterol regulation. Biochem. Soc. Trans. 52, 431–440. Alexa, V.T., Galuscan, A., Soica, C.M., Cozma, A., Coricovac, D., Borcan, F., Popescu, I., Mioc, A., Szuhanek, C. and Dehelean, C.A. 2022. In vitro assessment of the cytotoxic and antiproliferative profiles of bergamot, orange, and clove essential oil-containing natural preparations. Molecules 27, 990. Bairagee, D., Patel, P.S., Pradhan, J. and Panchawat, S. 2025. Cytotoxic effects of herbal medicine: an overview of mechanistic pathways and clinical relevance. Pharmacol. Res. Nat. Prod. 7, 100259. Bligh, E.G. and Dyer, W.J. 1959. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 37(8), 911–917. Chok, K.C., Ng, M.G., Ng, K.Y., Koh, R.Y. and Tiong, Y.L. 2021. Edible bird's nest: recent updates and laboratory-based industry insights. Front. Pharmacol. 12, 1–19. Cui, D., Yu, X., Guan, Q., Shen, Y., Liao, J., Liu, Y. and Su, Z. 2025. Cholesterol metabolism: molecular mechanisms, biological functions, diseases, and therapeutic targets. Mol. Biomed. 6(72), 6; doi:10.1016/j.mbm.2025.06.007 Duan, Y., Gong, K., Xu, S., Zhang, F., Meng, X. and Han, J. 2022. Regulation of cholesterol homeostasis in health and diseases: from mechanisms to targeted therapeutics. Signal Transduction Targeted Therapy 7, 265; doi:10.1016/j.sttt.2022.1265 Gan, J.Y., Chang, L.S., Mat, N.N.A., Babji, A.S. and Seng, J.L. 2020. Evaluation of the physicochemical properties, amino acid profile, and bioactivities of edible bird’s nest hydrolysate as affected by drying methods. LWT-Food. Sci. Technol. 131, 109777. Gavanji, S., Bakhtari, A., Famurewa, A.C. and Othman, E.M. 2023. Cytotoxic activity of herbal medicines as assessed in vitro: a review. Chem. Biodiversity 20, e202201098. Javitt, N.B. 1990. Hep G2 cells as a resource for metabolic studies: lipoprotein, cholesterol, and bile acids. FASEB. J. 4(2), 161–168. Kan, M., Ren, H., Du, J. and Sun, Y. 2025. The effect of hydrolysis on the bioactivity of edible bird’s nest: a systematic review. J. Future Foods; doi: 10.1016/j.jfutfo.2025.05.007 Lee, C.H., Lee, T.H., Wong, S.L., Nyakuma, B.B., Hamdan, N., Khoo, S.C., Ramachandran, H. and Jamaluddin, H. 2023. Characteristics and trends in global edible bird’s nest (EBN). J. Food. Meas. &. Characterization. 17(5), 4905–4926. Lee, T.H., Wani, W.A., Lee, C.H., Cheng, K.K. and Shreaz, S. 2021. Edible bird’s nest: functional values of prized animal-based bioproducts from Southeast Asia: a review. Front. Pharmacol. 12, 1–16. Li, X. and Li, M. 2024. Unlocking cholesterol metabolism in metabolic-associated steatotic liver disease: molecular targets and natural product interventions. Pharm 17, 1073. Ling, A.J.W., Chang, L.S., Babji, A.S., Latip, J., Koketsu, M. and Lim, S.J. 2022. Biochemistry, sources, extraction, and functions of sialic acid with special reference to edible bird’s nest. Food Chem. 367, 130755. Nasir, N. N. M., Ibrahim, R. M., Bakar, Z. A., Mahmud, R., and Razak, N. A. A. 2021. Influence of characterization and extraction on protein profiling of edible bird’s nest. Foods. 10, 2248. Nemudzivhadi, V. and Masoko, P. 2014. In vitro assessment of cytotoxicity, antioxidant, and anti-inflammatory activities of Ricinus communis (Euphorbiaceae) leaf extracts. Evid. Based Complement. Alternat. Med. 2014, 625961; doi:10.1155/2014/625961 Permatasari, H.K., Permatasari, Q.I., Taslim, N.A., Subali, D., Kurniawan, R., Surya, R., Qhabibi, F.R., Tanner, M.J., Batubara, S.C., Mayulu, N., Gunawan, W.B., Syauki, A.Y., Salindeho, N., Park, M.N., Lee, J.A.J., Tjancrawinata, R.R. and Kim, B. 2023. Eating bird nests as novel functional foods for combating metabolic syndrome: comprehensive in silico, in vitro, and in vivo studies. Nutrients 15, 3886. Pramfalk, C., Larsson, L., Härdfeldt, J., Eriksson, M. and Parini, P. 2016. Culturing of HepG2 cells with human serum improve their functionality and suitability in studies of lipid metabolism. Biochimica. Et. Biophysica. Acta. 1861(1), 51–59. Ramli, A.N.M., Badrulzaman, S.Z.S., Patil, R.V., Azelee, N.I.W., Manas, N.H.A. and Aminan, A.W. 2024. Beyond tradition: a novel approach for edible bird nest cleaning and its processing. Vet. Res. Commun. 48(1), 29–37. Rusanuar, N.D., Aminuddin, A., Hamid, A.A., Kumar, J., Hui, C.K., Mahadi, M.K. and Ugusman, A. 2025. The potential of edible bird’s nests in reducing cardiovascular disease risk factors: a narrative review. Int. J. Mol. Sci. 26(10), 4619. Shizhan, M., Sun, W., Gao, L. and Liu, S. 2019. Therapeutic targets of hypercholesterolemia: hMGCR and LDLR. Diabetes. Metabolic. Syndrome. Obesity. Targets. Therapy. 12, 1543–1553. Unal, K.I., Chang, L.S., Wan Mustapha, W.A., Mohd Razali, N.S., Babji, A.S. and Lim, S.J. 2022. Edible bird’s nest, a valuable glycoprotein source: current research prospects and challenges in Malaysia. Sains Malaysiana 51(9), 2829–2842; doi:10.1016/j.sainsmalaysia.2022 Verma, K., Makwana, S., Paliwal, S., Paliwal, V., Jain, S., Paliwal, S. and Sharma, S. 2022. Simvastatin ameliorates oxidative stress levels in HepG2 cells and hyperlipidemic rats. Curr. Res. Pharmacol. Drug Discov. 3, 100088. Yida, Z., H. Al-Shuwayah, M. Ismail, and M. U. Imam. 2022. The edible bird’s nest regulates hepatic cholesterol metabolism through the transcriptional regulation of cholesterol-related genes Evidence-based complementary and alternative medicine 8882993. Zulkefe, N.N., Ibrahim, M.A., Azuan, N.F., Ch’ng, S.E., Ismail, N., Bakar, M.Z.A. and Chan, K.W. 2024. A review of the quality and manufacturing standards of edible bird nests of the three largest exporting countries in the world. J. Food Qual. 5608357, 5608357./ | ||


| How to Cite this Article |
| Pubmed Style Sulaiman SZS, Noor MAM, Ilias AN, Titisari N, Fauzi A, Ajat MMM. Comparative effects of raw and commercialized edible bird’s nest products on low-density lipoprotein receptor expression and cholesterol regulation in HepG2 cells. Open Vet. J.. 2026; 16(5): 2883-2891. doi:10.5455/OVJ.2026.v16.i5.30 Web Style Sulaiman SZS, Noor MAM, Ilias AN, Titisari N, Fauzi A, Ajat MMM. Comparative effects of raw and commercialized edible bird’s nest products on low-density lipoprotein receptor expression and cholesterol regulation in HepG2 cells. https://www.openveterinaryjournal.com/?mno=297079 [Access: June 26, 2026]. doi:10.5455/OVJ.2026.v16.i5.30 AMA (American Medical Association) Style Sulaiman SZS, Noor MAM, Ilias AN, Titisari N, Fauzi A, Ajat MMM. Comparative effects of raw and commercialized edible bird’s nest products on low-density lipoprotein receptor expression and cholesterol regulation in HepG2 cells. Open Vet. J.. 2026; 16(5): 2883-2891. doi:10.5455/OVJ.2026.v16.i5.30 Vancouver/ICMJE Style Sulaiman SZS, Noor MAM, Ilias AN, Titisari N, Fauzi A, Ajat MMM. Comparative effects of raw and commercialized edible bird’s nest products on low-density lipoprotein receptor expression and cholesterol regulation in HepG2 cells. Open Vet. J.. (2026), [cited June 26, 2026]; 16(5): 2883-2891. doi:10.5455/OVJ.2026.v16.i5.30 Harvard Style Sulaiman, S. Z. S., Noor, . M. A. M., Ilias, . A. N., Titisari, . N., Fauzi, . A. & Ajat, . M. M. M. (2026) Comparative effects of raw and commercialized edible bird’s nest products on low-density lipoprotein receptor expression and cholesterol regulation in HepG2 cells. Open Vet. J., 16 (5), 2883-2891. doi:10.5455/OVJ.2026.v16.i5.30 Turabian Style Sulaiman, Sharifah Zakiah Syed, Mohd Akmal Mohd Noor, Amirul Nazhan Ilias, Nurina Titisari, Ahmad Fauzi, and Mohd Mokrish Mohd Ajat. 2026. Comparative effects of raw and commercialized edible bird’s nest products on low-density lipoprotein receptor expression and cholesterol regulation in HepG2 cells. Open Veterinary Journal, 16 (5), 2883-2891. doi:10.5455/OVJ.2026.v16.i5.30 Chicago Style Sulaiman, Sharifah Zakiah Syed, Mohd Akmal Mohd Noor, Amirul Nazhan Ilias, Nurina Titisari, Ahmad Fauzi, and Mohd Mokrish Mohd Ajat. "Comparative effects of raw and commercialized edible bird’s nest products on low-density lipoprotein receptor expression and cholesterol regulation in HepG2 cells." Open Veterinary Journal 16 (2026), 2883-2891. doi:10.5455/OVJ.2026.v16.i5.30 MLA (The Modern Language Association) Style Sulaiman, Sharifah Zakiah Syed, Mohd Akmal Mohd Noor, Amirul Nazhan Ilias, Nurina Titisari, Ahmad Fauzi, and Mohd Mokrish Mohd Ajat. "Comparative effects of raw and commercialized edible bird’s nest products on low-density lipoprotein receptor expression and cholesterol regulation in HepG2 cells." Open Veterinary Journal 16.5 (2026), 2883-2891. Print. doi:10.5455/OVJ.2026.v16.i5.30 APA (American Psychological Association) Style Sulaiman, S. Z. S., Noor, . M. A. M., Ilias, . A. N., Titisari, . N., Fauzi, . A. & Ajat, . M. M. M. (2026) Comparative effects of raw and commercialized edible bird’s nest products on low-density lipoprotein receptor expression and cholesterol regulation in HepG2 cells. Open Veterinary Journal, 16 (5), 2883-2891. doi:10.5455/OVJ.2026.v16.i5.30 |

