Open Veterinary Journal, (2026), Vol. 16(5): 2860-2868
Research Article
10.5455/OVJ.2026.v16.i5.28
Lactiplantibacillus plantarum-fermented shallot (Allium cepa L.) extract: A novel natural antibiotic alternative in drinking water for Escherichia coli-challenged broilers
Hoang Thi Anh Phuong1,3, Nguyen Dinh Vinh2, Nguyen Xuan Hoa3, Vo Thi Minh Tam3 and
Phan Vu Hai3*
1Department of Veterinary Medicine, Tay Nguyen University, Buon Ma Thuot City, Vietnam
2School of Agriculture and Natural Resources, Vinh University, Vinh City, Vietnam
3Faculty of Animal Husbandry and Veterinary Medicine, University of Agriculture and Forestry, Hue University, Hue City, Vietnam
*Corresponding Author: Phan Vu Hai. Faculty of Animal Husbandry and Veterinary Medicine, University of Agriculture and Forestry, Hue University, Hue City, Vietnam. Email: phanvuhai [at] hueuni.edu.vn
Submitted: 13/11/2025 Revised: 11/03/2026 Accepted: 21/03/2026 Published: 31/05/2026
© 2025 Open Veterinary Journal
This is an Open Access article distributed under the terms of the Creative Commons Attribution-Non Commercial-No Derivatives License (http://creativecommons.org/licenses/by-nc-nd/4.0/), which permits non-commercial re-use, distribution, and reproduction in any medium, provided the original work is properly cited, and is not altered, transformed, or built upon in any way.
ABSTRACT
Background: The global poultry industry is under pressure to reduce its reliance on antibiotics, necessitating the search for effective alternatives. Pathogenic Escherichia coli infections remain a major challenge, leading to significant economic losses due to impaired growth performance and increased mortality.
Aim: This study aimed to evaluate the efficacy of shallot extract (Allium cepa L.) fermented by Lactiplantibacillus plantarum 1582 fermented shallot (FS) as a potential antibiotic alternative in experimentally challenged E. coli broiler chickens.
Methods: A total of 300 1-day-old J-Dabaco broilers were randomized to six treatments (n=50; five replicates of 10): negative control (NC), (PC), antibiotic control (AB), and three challenged groups receiving FS at 1%, 2%, or 3% (FS1–FS3). Challenged birds were orally inoculated with E. coli ExPEC_A338 (3 × 108 CFU/ml) on days 7 and 8. Outcomes included growth performance, cecal microbiota, jejunal histomorphology, immune-organ indices, serum Immunoglobulin M (IgM), Immunoglobulin A (IgA), and Immunoglobulin G levels, and mRNA expression of tight-junction proteins (ZO-1, occludin, and claudin-2) and cytokines [IL-4, IL-1β, TNF-α, and Interferon-gamma (IFN-γ)].
Results: Supplementation with FS, especially the 3% dose (FS3), significantly improved body weight gain by 9.4% and restored survival to 100%, matching the NC and AB groups. FS3 enhanced gut health by reducing the densities of pathogenic E. coli and Salmonella spp. while simultaneously increasing the densities of beneficial Lactobacillus spp. (p < 0.05). Morphological analysis showed that the FS3 group had the highest villus height (VH) (1,049.86 µm) and VH/Crypt Depth ratio (13.05) (p < 0.05). Although immune-organ indices remained unchanged, dietary FS at 2%–3% significantly upregulated ZO-1, while IFN-γ increased at 1% FS (FS1) but remained comparable to the challenged control at 2%–3% (p < 0.05). Serum IgM increased at 1%–2%, and serum IgA decreased at 3% (p < 0.05).
Conclusion: Collectively, these findings demonstrate that L. plantarum-FS extract, particularly at a 3% inclusion level, represents a promising non-antibiotic strategy for broilers challenged with E. coli, primarily through enhanced growth performance, intestinal integrity, and a favorable gut microbial balance.
Keywords: Antibiotic alternative, Broiler, Escherichia coli, Fermentation, Lactiplantibacillus plantarum.
Introduction
Avian pathogenic Escherichia coli (APEC) is one of the most critical bacterial agents impacting the poultry industry, causing significant global economic losses through systemic lesions such as polyserositis and cellulitis (Mehat et al., 2021). This challenge is further exacerbated by the zoonotic potential of contaminated poultry products (Elsharawy, 2022) and the rapid escalation of antimicrobial resistance, emphasizing the urgent need for effective alternatives to conventional antibiotics (Sebastian et al., 2021).
Phytogenic substances, particularly those derived from the Allium genus, have garnered attention owing to their potent bioactive components, such as quercetin and allicin (Salehi et al., 2019; Hai et al., 2025a). The antimicrobial and immunomodulatory activities of these compounds have been documented (Hai and Hoa, 2020; Hai et al., 2025b). However, their efficacy is often constrained by poor bioavailability, as many active components remain trapped within the plant cell wall matrix (Liu, 2013).
Fermentation-targeted bioprocessing offers a rational strategy to disrupt plant cell architecture, liberate bound phytochemicals, and potentiate their biological activity (Gao et al., 2023). Although spontaneous fermentation can yield inconsistent outcomes and raise safety concerns (Chen et al., 2018), controlled fermentation with lactic acid bacteria (LAB) provides a safer and reproducible platform. LAB, particularly Lactiplantibacillus spp., can competitively inhibit enteric pathogens, such as E. coli, and modulate host immune function (De Montijo-Prieto et al., 2023). Nevertheless, probiotics alone have shown limited protection under severe APEC challenge, suggesting that single-modality interventions may be insufficient (Rahayu et al., 2021). The putative synergy of LAB-fermented botanical extracts, which combine antimicrobial phytochemicals with probiotic and postbiotic effects, remains insufficiently characterized in E. coli challenge models.
To address this gap, this study evaluated the effects of a Lactiplantibacillus plantarum-fermented shallot (FS) (Allium cepa L.) extract administered via drinking water on growth performance, gut health, and immune responses in broiler chickens under experimental E. coli challenge. We hypothesized that the fermented extract would mitigate infection-associated performance losses by reinforcing intestinal barrier integrity, reshaping the gut microbiota toward beneficial taxa, and modulating systemic and mucosal immunity.
Materials and Methods
FS extract
Shallots (A. cepa L., GenBank ID: NC_057575.1) meeting the Vietnamese Good Agricultural Practices standards were purchased from Dien Mon, Thua Thien Hue, Vietnam. The raw material was pretreated (washed, peeled, and soaked in a 5% NaCl solution for 120 minutes) and then homogenized.
Microbial strain and fermentation process
The strain L. plantarum 1582 (GenBank ID: MT597487.1), isolated from indigenous chicken, was selected based on its resistance to E. coli and shallot extract (Phuong et al., 2024). The strain was activated in de Man, Rogosa, and Sharpe (MRS) broth (Oxoid, UK) at 37℃ for 24 hours.
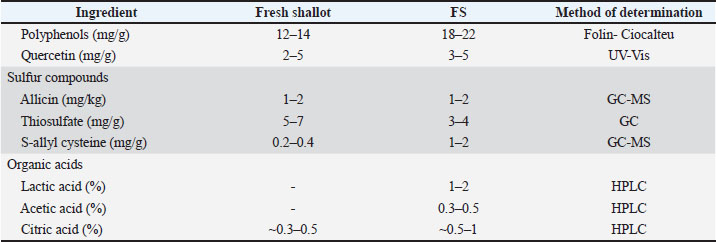
The fermentation process followed the method described by Hai et al. (2025a), with modifications. The homogenized shallot was inoculated with a 1% inoculum ratio of L. plantarum 1582 (108 CFU/ml) in a medium supplemented with 4% NaCl and 3% glucose. The mixture was anaerobically incubated (Kuvings KGC-712CB) at 37℃ at 60 rpm for 72 hours. The extract was filtered to yield the final product (~2.7–3 × 107 CFU/ml) and stored at 4℃. Table 1 presents the main bioactive components.
Table 1. Bioactive constituents of fresh and L. plantarum-FS (A. cepa L.) extracts.
Pathogenic bacterial strains
The virulent E. coli strain ExPEC_A338 (GenBank ID: CP142559.1) (Hai et al., 2015c) was activated in Luria–Bertani broth, centrifuged, washed, and diluted in phosphate-buffered saline (PBS) buffer (pH 7.2) to the required concentration.
Basal diet
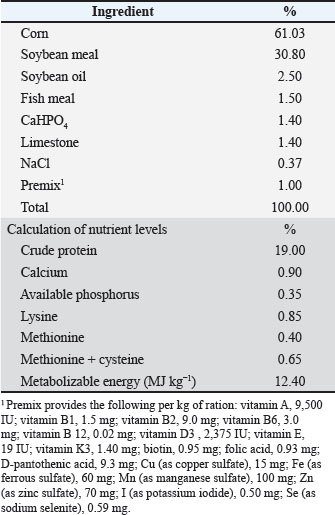
Birds were fed a basal diet (Table 2) based on corn and soybean meal, which met the feeding standards for broilers set by the Ministry of Agriculture and Rural Development of Vietnam (TCN 661-2005) and was antibiotic-free.
Table 2. The nutritional composition of the basic diet.
Animals used and experimental design
A total of 300 1-day-old J-Dabaco colored feather male broilers were brooded communally (Days 1–7) and subsequently randomly allocated in a completely randomized design: 6 treatments × 5 replicate pens × 10 birds/pen: NC (negative control: no supplement, no challenge; isolated rearing), PC (positive control: challenged), FS1–FS3 (supplemented with 1%–3% FS in drinking water daily, challenged), and AB (antibiotic: Terra-Neocine - Mebipha JSC - Vietnam at 1 g/l for 5 days pre-challenge and 2 g/l for 5 days post-challenge).
On days 7 and 8, the birds in treatments (2) through (6) were challenged via oral gavage with 0.5 ml of E. coli ExPEC_A338 suspension (3 × 108 CFU/ml). The NC group received 0.5-ml of PBS. Figure 1 summarizes the experimental layout and evaluated parameters.

Fig. 1. Flowchart of the experimental design and parameters evaluated.
All procedures for the care, housing, and slaughter of the experimental chickens complied with standards approved by the Animal Ethics Advisory Committee of Hue University, Vietnam.
Data collection and parameter assessment
Growth performance
Feed intake (FI) for each pen was recorded daily at 07:00 hours, and body weight (BW) was measured at the beginning and end of the experiment. Mortality was checked daily to determine the survival rate. From these raw data, body weight gain (BWG, end BW - initial BW) and feed conversion ration (FCR, total FI/BWG) were calculated. The production efficiency index (PEI) was calculated using the following formula: (BWG (kg) × Survival rate (%) × 100)/(FCR × Experiment days) (Martins et al., 2016).
Sample collection
On day 35, 15 birds/pen were randomly selected, anesthetized, and sampled. Blood was centrifuged at 3,000 rpm for 10 minutes to obtain serum. Immune organs (bursa of fabricius, thymus, and spleen) were weighed to calculate relative organ weights (g/100 g BW). Cecal digesta samples were aseptically collected for microbial analysis. Ileal tissue (midsection) was collected for histological and gene expression analyses.
Immunoglobulin content
Serum was isolated from blood samples for 10 minutes and subsequently preserved at −20°C until analysis. Serum immunoglobulin levels, including IgA (MBS705241), IgM (MBS706158), and Immunoglobulin G (IgG) (MBS260043), were quantified using commercial enzyme-linked immunosorbent assay kits (MyBioSource, San Diego, CA). The absorbance was read at 450 nm, and the final concentrations were derived from the respective standard curves.
Gene expression analysis
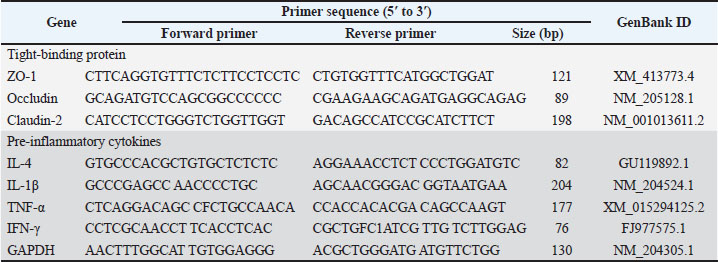
Total RNA was extracted from the ileal mucosa (Trizol, Invitrogen), and cDNA was synthesized (FIRE Script RT cDNA, Solid Biodyne). Reverse transcription quantitative polymerase chain reaction (RT-qPCR) was performed on a QuantStudio™ 5 system (Thermo Fisher) to quantify the relative expression of genes related to immunity [IL-4, IL-1β, TNFα, Interferon-gamma (IFNγ)] and tight junction proteins (ZO-1, claudin-2, and occludin). Gene expression was calculated using the 2⁻ΔΔCt method (Livak and Schmittgen, 2001) with glyceraldehyde 3-phosphate dehydrogenase (GAPDH) as the reference gene (Table 3).
Table 3. Primer sequences used for RT-qPCR.
Gut health
The population of E. coli, Salmonella spp., and Lactiplantibacillus spp. in cecal digesta was quantified by plate counting on selective agar media: Eosin Methylene Blue Agar supplemented with 1% CaCO3, Salmonella Shigella agar, and MRS agar. The corresponding reference standards were ISO 13349/2001, ISO 6579/2003, ISO 7937/2004, and ISO/Dis 11290/1994. The results were transformed into log10 CFU/g.
Histological morphology
Ileal samples were fixed in 10% formalin, processed histologically (5 μm sections), and stained with Hematoxylin and Eosin (H&E) (Liu et al., 2025). Villus height (VH), crypt depth (CD), and surface area were measured using Image-Pro Plus software (v. 6.0).
Statistical analyses
Data were analyzed using the SPSS 22.0 software. The pen was considered the experimental unit for growth performance (n=5 pens per treatment). Three birds per pen were sampled for microbiology, histomorphology, immune-organ indices, serum immunoglobulins, and gene expression data (n=15 birds per treatment). Treatment effects were evaluated using the general linear model with treatment as a fixed effect. Mean separation was performed using one-way analysis of variance with Bonferroni post hoc test. The chi-square test (χ²) was used for the frequency data (survival rate). Differences were considered statistically significant at p < 0.05. Data are presented as Mean ± Standard Error of the mean.
Ethical approval
All procedures concerning the care, housing, and slaughter of the experimental chickens were carried out in accordance with standards approved by the Animal Ethics Advisory Committee, Hue University, Vietnam (March 20, 2024).
Results
Effects of FS extract on broiler performance
The average water intake was monitored daily; the mean intake per bird was 135–185 ml/day (days 8–35). Based on the viable count of the FS (~2.7–3.0 × 10⁷ CFU/ml), the estimated daily intakes of viable L. plantarum 1582 were approximately 3.6–5.6 × 10⁷, 7.2–11.1 × 10⁷, and 1.1–1.7 × 10⁸ CFU/bird/day for the 1%, 2%, and 3% supplementation levels, respectively. During the pre-challenge period (days 1–7), BWG, FI, FCR, and survival did not differ among treatments (p=0.26), indicating comparable baseline performance (Table 4). Following E. coli challenge (days 3–5), the challenged control (PC) exhibited marked performance depression, with BWG, FI, and survival reduced to 593.57 g, 1,030.10 g, and 76.67%, respectively. Moreover, characteristic APEC lesions were observed in the deceased birds (Fig. 2). Supplementation with FS extract mitigated these losses. Relative to PC, BWG increased by +3.05%, +8.15%, and +8.54% in FS1, FS2, and FS3, respectively, and survival was restored to 93.33%–100%, comparable to the NC and AB (Table 4).
Table 4. Effect of FS supplementation on broiler chicken growth performance.
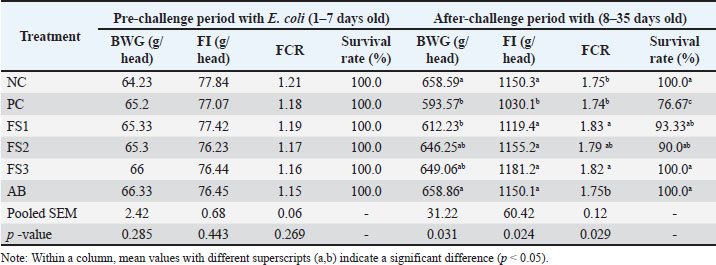
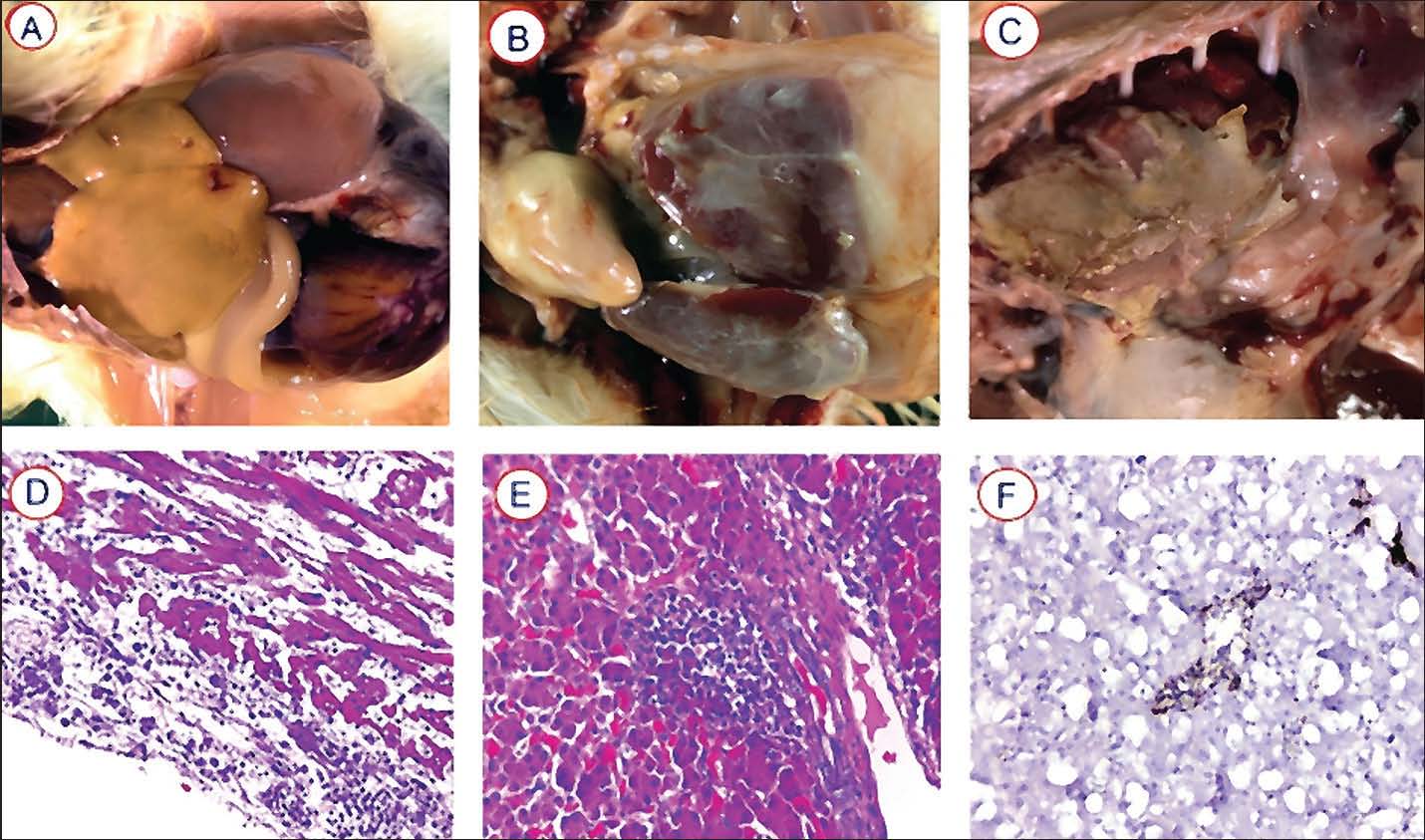
Fig. 2. Chickens, post-E. coli challenge, exhibited gross pathological lesions: (A) Distended, congested yolk sac with prominent blood vessels, containing abnormal brown fluid; (B) Polyserositis (comprising pericarditis, perihepatitis, peritonitis, and airsacculitis) resulting from the systemic spread of E. coli; (C) Pleuropneumonia and airsacculitis in broilers; and histopathological lesions observed under microscopy (H&E, 40×): (D) Liver with multiple areas of scattered fibrin deposition; (E) Pericardium characterized by an infiltration of intact or degenerated heterophils, macrophages, lymphocytes, and plasma cells, along with fibrin; (F) Fibrinoid necrosis was also evident in the follicular centers and blood vessels of the spleen, particularly the splenic arterioles.
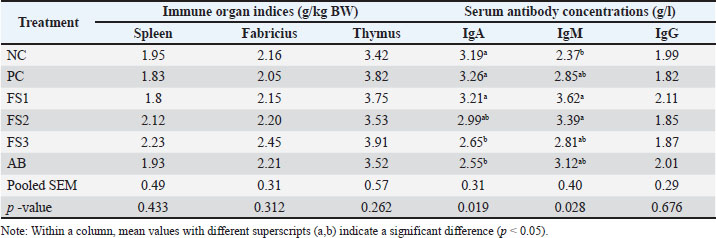
Effects of FS extract on immune function
Table 5 shows the effects of treatments on immune organ indices and serum antibodies. No significant differences were observed in the relative weights of the spleen, bursa of Fabricius, or thymus (p > 0.05) or serum IgG concentration (p > 0.05). However, dietary treatment significantly modulated serum immunoglobulins: IgA differed among groups (p = 0.019), being lower in FS3 and AB than in NC, PC, and FS1; IgM also varied (p = 0.028), with higher concentrations in FS1 and FS2 than in NC.
Table 5. Immune organ indices and serum antibody concentrations of experimental chickens.
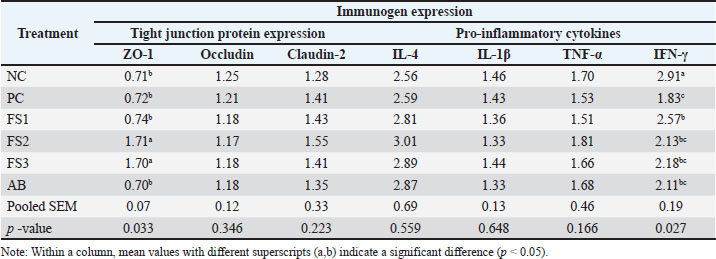
Tight-junction gene expression was modulated by the dose of FS. ZO-1 mRNA abundance was significantly higher in FS2 (1.71) and FS3 (1.70) than in PC (0.72) (p < 0.05). No significant differences in ZO-1 were detected among the PC, NC, FS1, and AB groups (p > 0.05). IFN-γ expression also differed among treatments (Table 6): NC displayed the highest level (2.91), significantly exceeding all other groups, whereas PC had the lowest (1.83). FS1 (2.57) was significantly higher than PC (p < 0.05), whereas FS2 (2.13), FS3 (2.18), and AB (2.11) did not differ from PC but were lower than NC.
Table 6. Effects of FS supplementation on the expression of immune genes.
Effects of FS extract on gastrointestinal health
FS altered the cecal microbiology and ileal histomorphology (Table 7). Representative images of ileal mucosal morphology are shown in Figure 3. Pathogen loads were greatest in PC (Salmonella spp., 6.67 log CFU/g; E. coli, 7.01 log CFU/g). Both FS and AB reduced these counts (p < 0.05), with FS3 exhibiting the lowest levels (Salmonella, 4.20 log CFU/g; E. coli, 5.20 log CFU/g), equal to or lower than AB (4.63 and 5.63 log CFU/g, respectively). The number of beneficial bacteria increased with FS, and FS3 achieved the highest Lactobacillus spp. density (7.21 log CFU/g), surpassing NC (4.83), PC (5.48), and AB (4.32) (p < 0.05).
Table 7. Effect of fermented supplement on intestinal health.
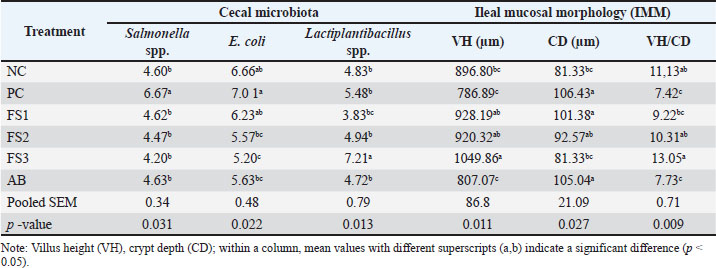
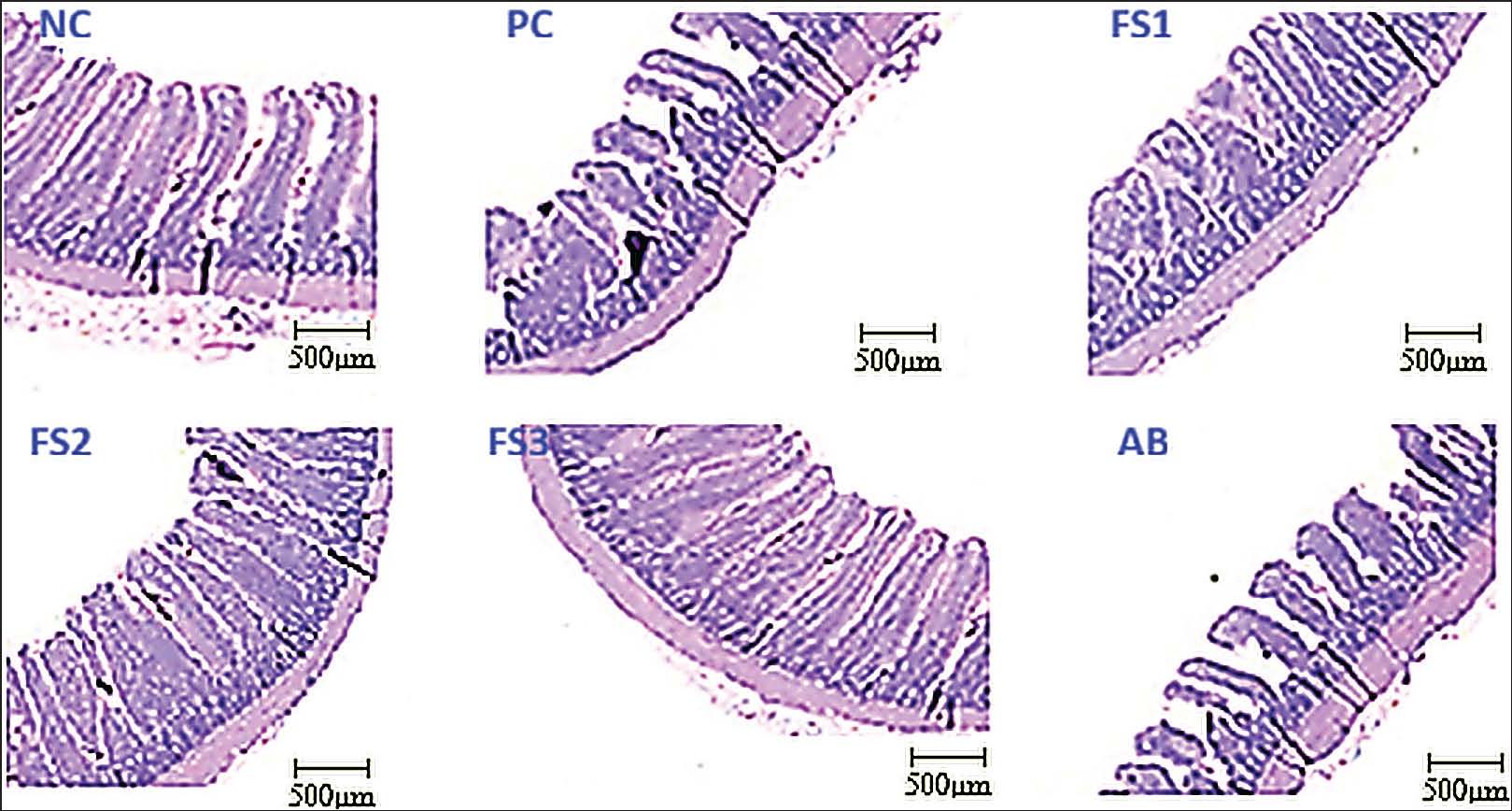
Fig. 3. Representative images of ileal mucosal lesions in E. coli-infected chickens from the positive control (PC) and antibiotic (AB) groups, which were ameliorated in the FS2 group and particularly in the FS3 group following FS supplementation.
Consistent with microbial shifts, ileal morphology improved in the FS group (p < 0.05). VH was greatest in FS3 (1,049.86 μm), exceeding NC (896.80 μm), PC (786.89 μm), and AB (807.07 μm); FS1 and FS2 were also higher than these three groups. CD in FS3 (81.33 μm) was similar to that in NC but significantly lower than that in PC, AB, FS1, and FS2 (92.57–106.43 μm). Consequently, the VH:CD ratio was the highest in FS3 (13.05), significantly exceeding PC, AB, and FS1 (7.42–9.22). Collectively, these results indicate the dose-responsive benefits of FS on performance, immune readouts, and gut health under E. coli challenge.
Discussion
Effects of FS extract on broiler performance
Growth improvements in FS birds are biologically plausible. Shallot contains antioxidant, anti-inflammatory, and antimicrobial polyphenols, flavonoids, and organosulfur compounds (Table 1). These compounds likely inhibited E. coli proliferation, improved intestinal homeostasis, and enhanced nutrient use. Fermentation increases efficacy by breaking plant cell walls, boosting bioavailability, and producing short-chain fatty acids that support enterocytes. Functionally, FS acts as a synbiotic. It combines prebiotic substrates from shallot with L. plantarum’s probiotic effects to lower luminal pH and suppresses pathogens.
These findings agree with those of previous studies. Fermented botanicals improve broiler performance (Hai et al., 2020; Adli et al., 2024), and lactic acid bacteria enhance growth and feed efficiency (Liu et al., 2025). Although the FCR was unchanged, the PEI increased in FS2 and FS3 vs the challenged control. This shows net production gains when morbidity and mortality are considered. FS offers a promising antibiotic-sparing strategy at 3% inclusion.
Effects of FS extract on immune function
No changes appeared in the spleen, bursa of Fabricius, or thymus indices. This indicates no overt lymphoid hypertrophy or atrophy. Serum immunoglobulins showed dose-dependent modulation. Elevated IgM at 1%–2% may reflect an early antibody response. Lower IgA at 3% (similar to the antibiotic group) likely indicates reduced antigenic load from a stronger barrier, although direct suppression cannot be ruled out (Ghareeb and K, 2012), but it may also reflect changes in mucosal immune activity. Therefore, we cautiously interpreted the IgA result and recommend future studies measuring mucosal secretory IgA and local immune-cell responses.
ZO-1 increased in FS2 and FS3, indicating tight-junction architecture reinforcement. ZO-1 is a central scaffolding protein that links transmembrane indicating tight junction components to the actin cytoskeleton (Gonzalez-Mariscal et al., 2003). FS mainly strengthens the junctional scaffold (Awad et al., 2009), matching the anti-inflammatory effects of fermented Allium compounds (Arreola et al., 2015).
E. coli challenge lowered IFN-γ expression. FS at 1% partially restored it toward non-challenged levels. However, 2%–3% doses remained similar to the challenged control. This may reflect a U-shaped dose-response or lower antigenic stimulation (Marefati et al., 2021). The absence of significant effects occurred on IL-1β or TNF-α. Overall, FS acts as a dose-dependent immunomodulator that concurrently strengthens epithelial defenses.
Effects of FS extract on gastrointestinal health
These results indicate that FS effectively modulates the intestinal microbiota. At 3% (FS3), it suppressed Salmonella spp. and E. coli to levels equal to or better than the antibiotic group (Mountzouris et al., 2010; Gaggìa et al., 2010). FS3 also markedly increased beneficial Lactobacillus spp. markedly—an effect absent in the antibiotic group. Lactobacillus spp. protect the mucosa by competing for adhesion sites and producing bacteriocins (Lee et al., 2011). Ileal VH increased with FS, especially at 3% (FS3) (Awad et al., 2009), whereas CD decreased to nonchallenged levels in FS3 (Pelicano et al., 2005). Thus, the VH:CD ratio was the highest in FS3, surpassing that in both the control and antibiotic groups.
Conclusion
Supplementation of drinking water with 3% L. plantarum- FSextract effectively mitigated E. coli challenge in broilers. The 3% FS group achieved BWG, PEI, and 100% survival, which were comparable to those of the nonchallenged and antibiotic controls and superior to those of the challenged control. Benefits aligned with improved gut health—suppressed cecal pathogens, enrichment of Lactobacillus spp., and optimized ileal histomorphology (higher VH, reduced CD, greater VH:CD) —supporting absorption and mucosal stability. FS displayed dose-dependent immunomodulation: 1%–2% increased serum IgM and 1% increased IFN-γ expression, whereas 3% decreased serum IgA and upregulated ZO-1. The absence of a non-FS extract control was a limitation; future studies should directly compare FS and non-fermented extract to confirm fermentation-specific effects.
Acknowledgments
We thank laboratory staff for technical assistance and animal care during the study.
Conflict of interest
The authors declare no conflicts of interest.
Funding
This project was supported by the Ministry of Education and Training, Vietnam (grant no. B2023-DHH-24).
Authors’ contributions
Experiment design: P.V.H., N.D.V., N.X.H., and H.T.A.P; Data analysis: P.V.H. and H.T.A.P.; Administrative and technical support: N.D.V. and V.T.M.T.; and manuscript drafting: P.V.H. and H.T.A.P.
Data availability
Data are stored according to the project records. The findings are provided by the corresponding author upon request.
References
Adli, D.N., Sholikin, M.M., Ujilestari, T., Ahmed, B., Sadiqqua, A., Harahap, M.A., Sofyan, A. and Sugiharto, S. 2024. Effect of fermentation of herbal products on growth performance, breast meat quality, and intestinal morphology of broiler chickens: a meta-analysis. Ital. J. Anim. Sci. 23, 734–750.
Arreola , R., Quintero-Fabián, S., López-Roa, R.I., Flores-Gutiérrez, E.O., Reyes-Grajeda, J.P., Carrera-Quintanar, L. and Ortuño-Sahagún, D. 2015. Immunomodulation and anti-inflammatory effects of garlic compounds. J. Immunol. Res. 2015, 401630.
Awad, W.A., Ghareeb, K., Abdel-Raheem, S. and Böhm, J. 2009. Effects of dietary inclusion of probiotic and synbiotic on growth performance, organ weights, and intestinal histomorphology of broiler chickens. Poult. Sci. 88, 49–56.
Chen, X., Zhang, Y., Wang, Z., Li, J., Zhang, H. and Li, Y. 2018. Hepatoprotective effects of Lactobacillus on carbon tetrachloride-induced acute liver injury in mice. Int. J. Food Sci. Nutr. 69, 1–10.
De Montijo-prieto, S., Razola-Díaz, M.D.C., Barbieri, F., Tabanelli, G., Gardini, F., Jiménez-Valera, M., Ruiz-Bravo, A., Verardo, V. and Gómez-Caravaca, A.M. 2023. Impact of lactic acid bacteria fermentation on phenolic compounds and antioxidant activity of avocado leaf extracts. Antioxidants 12, 298.
Elsharawy, N.T. 2022. Escherichia coli O157: h7 in meat and poultry: transmission, consequences on human health, and impact of non-thermal decontamination technologies: a review. Res. J. Food. Sci. Nutr. 7, 1–10.
Gaggìa, F., Mattarelli, P. and Biavati, B. 2010. Probiotics and prebiotics in animal feeding for safe food production. Int. J. Food Microbiol. 141, S15–S28.
Gao, Y., Li, X. and Wang, Y. 2023. Advancing herbal medicine: enhancing product quality and safety through fermentation. Front. Microbiol. 14, 991029.
Gonzalez-Mariscal, L., Betanzos, A., Nava, P. and Jaramillo, B.E. 2003. Tight junction proteins. Prog. Biophys. Mol. Biol. 81, 1–44.
Hai, P.V., Anh, L.X. and Hoa, N.X. 2025a. Fermented chive (Allium schoenoprasum) with Lactobacillus plantarum: a potential antibiotic alternative feed additive for broilers challenged with Escherichia coli. Fermentation 11, 277.
Hai, P.V., Dung, H.T., Thao, T.N., Hoa, N.X. and Hung, P.H.S. 2025b. Enhanced bioactivity and anti-microbial properties of Lactobacillus plantarum-fermented purple onion (Allium cepa L.) extracts against selected poultry microbes. Poult. Sci. J. 13, 159–169.
Hai, P.V., Hoa, N.X. and Phuong, H.T.A. 2025c. Fermented purple onion (Allium cepa L.) and chive (Allium schoenoprasum) bulb extracts as antibiotic alternatives against toxin-carrying bacteria: in vitro and pathogenicity assessment in chickens. Open. Vet. J. 15, 2355–2364.
Hai, P.V., Hung, P.H.S., Dung, H.T., Liem, T.N., Khuong, N.D.T. and Hoa, N.X. 2020. The dietary supplement efficiency of essential oil of chive (Allium macrostemon) on the productivity and health performance of broilers. Can Tho Univ. J. Sci. 12(1), 1–6.
Hai, P.V. and Hoa, N.X. 2020. Effect of Allium schoenoprasum extract on immune status against Newcastle virus and growth performance of broiler chicken. Hue Univ. J. Sci. Nat. Sci. 4, 2058–2064.
Lee, H., Yoon, H., Ji, Y., Kim, H., Park, H., Lee, J., Shin, H. and Holzapfel, W.H. 2011. Functional properties of Lactobacillus strains isolated from kimchi. Int. J. Food Microbiol. 145, 155–161.
Liu, R.H. 2013. Dietary bioactive compounds and their health implications. J. Food Sci. 78 Suppl 1, A18–A25.
Liu, Y., Wang, Z., Xi, W., Yuan, J., Zhang, K., Liu, H., Zhao, J. and Wang, Y. 2025. Lactiplantibacillus plantarum improves the growth performance and meat quality of broilers by regulating the cecal microbiota and metabolites. Front. Microbiol. 16, 1519552.
Livak, K.J. and Schmittgen, T.D. 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-ΔΔC(T)) method. Methods 25, 402–408.
Marefati, N., Ghorani, V., Shakeri, F., Boskabady, M., Kianian, F., Rezaee, R. and Boskabady, M.H. 2021. Anti-inflammatory, antioxidant, and immunomodulatory effects of Allium cepa and its main constituents. Pharm. Biol. 59, 287–302.
Martins, J.M.S., Carvalho, C.M.C., Litz, F.H., Silveira, M.M., Moraes, C.A., Silva, M.C.A., Fagundes, N.S. and Fernandes, E.A. 2016. Productive and economic performance of broiler chickens subjected to different nutritional plans. Rev. Bras. Zootec. 45, 399–407.
Mehat, J.W., Van Vliet, A.H.M. and La Ragione, R.M. 2021. The avian pathogenic Escherichia coli (APEC) pathotype is comprised of multiple distinct, independent genotypes. Avian Pathol. 50, 402–416.
Mountzouris, K.C., Tsitrsikos, P., Kalamara, E., Nitsch, S., Schatzmayr, G. and Fegeros, K. 2010. Evaluation of a probiotic containing Lactobacillus, Bifidobacterium, Enterococcus and Pediococcus strains in promoting broiler performance and modulating cecal microflora composition and metabolic activities. Poult. Sci. 89, 58–67.
Pelicano, E.R.L., de Souza, P.A., de Souza, H.B.A., Figueiredo, D.F., Boiago, M.M., Carvalho, S.R. and Zeola, N.M.B.L. 2005. Intestinal mucosa development in broiler chickens fed natural growth promoters. Braz. J. Poult. Sci. 7(4), 221–229.
Phuong, H.T.A., Hai, P.V., Giang, L.T.K., Duc, L.M., Khuong, N.D.T. and Hoa, N.X. 2024. Isolation and selection of potential probiotic Lactobacillus strains resistant to some Gram-negative bacteria causing diarrhea in chicken. Hue Univ. J. Sci. Agric. Rural Dev. 133, 117–134.
Rahayu, E.S., Utami, T. and Wulandari, L. 2021. Herbs as antibiotic alternatives in poultry: a review. Plants 10, 1804.
Salehi, B., Venditti, A., Frezza, C., Yücetepe, A., Altuntaş, Ü., Uluata, S., Butnariu, M., Sarac, I., Shaheen, S., Petropoulos, S.A., Matthews, K.R., Kılıç, C.S., Atanassova, M., Adetunji, C.O., Ademiluyi, A.O., Özçelik, B., Tsouh Fokou, P.V., Martins, N., Cho, W.C. and Sharifi-Rad, J. 2019. Apium plants: Beyond simple food and phytopharmacological applications. Appl. Sci. 9(17), 3547.
Sebastian, S., Tom, A.A., Babu, J.A. and Joshy, M. 2021. Antibiotic resistance in Escherichia coli isolates from poultry environment and UTI patients in Kerala, India: a comparison study. Comparative Immunol. Microbiol. Infect. Dis. 75, 101614.
Sharifi-Rad, M., Varoni, E.M., Salehi, B., Sharifi-Rad, J., Matthews, K.R., Ayatollahi, S.A., Kobarfard, F., Ibrahim, S.A., Mnayer, D., Zakaria, Z.A., Sharifi-Rad, M., Yousaf, Z., Iriti, M., Basile, A. and Rigano, D. 2020. Plants of the genus Zingiber as a source of bioactive phytochemicals: from tradition to pharmacy. Molecules 25, 4021.






