




| Research Article | ||
Open Vet. J.. 2026; 16(1): 241-255 Open Veterinary Journal, (2026), Vol. 16(1): 241-255 Research Article Effect of substituting fishmeal with black soldier fly and mealworm larvae meal on nutrient digestibility, body and mineral composition of Labeo rohitaNida Ismat1*, Sikandar Hayat1 and Majid Hussain21Institute of Molecular Biology and Biotechnology, The University of Lahore, Lahore, Pakistan 2Department of Fisheries and Aquaculture, University of Okara, Okara, Pakistan *Corresponding Author: Nida Ismat. Institute of Molecular Biology and Biotechnology, The University of Lahore, Lahore, Pakistan. Email: nidacheema7 [at] gmail.com Submitted: 28/08/2025 Revised: 25/11/2025 Accepted: 07/12/2025 Published: 31/01/2026 © 2025 Open Veterinary Journal
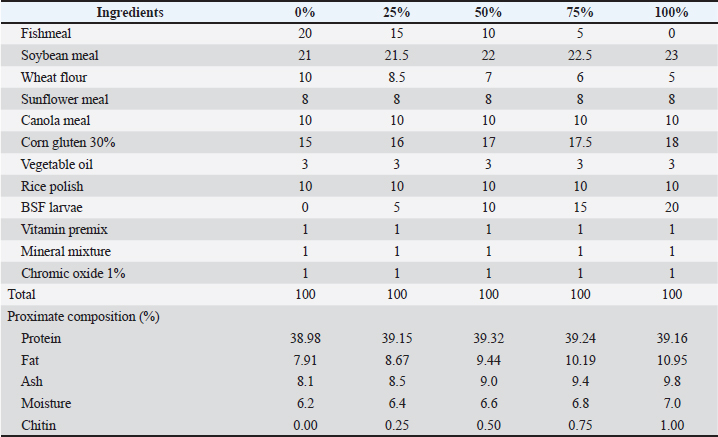
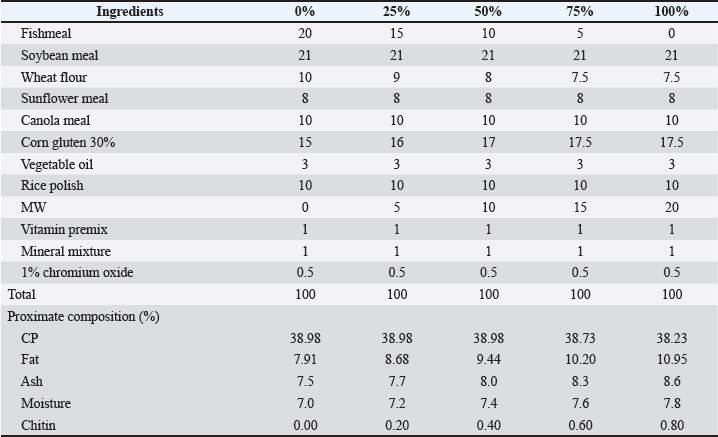
AbstractBackground: Aquaculture is a cornerstone of global food security, offering an essential source of protein and nutrition to an ever-growing human population. Fishmeal is a high-protein feed ingredient typically made from wild fish, which increases the depletion of wild fish. Insects such as black soldier fly (BSF) (Hermetia illucens) and mealworm (MW) (Tenebrio molitor) larvae meal are promising alternative protein sources for aquafeeds. However, their effects on Labeo rohita remain underexplored. Aim: This study evaluated the impact of substituting fishmeal with BSF and MW larvae meal on the nutrient digestibility and body and mineral composition of L. rohita fingerlings. Methods: A 60-day feeding trial was conducted using varying levels (0%, 25%, 50%, 75%, and 100%) of BSF and MW diets. Growth performance was assessed using the following parameters: weight gain (WG), feed conversion ratio (FCR), and specific growth rate (SGR). Digestibility trials measured apparent digestibility coefficients for protein, lipid, and gross energy. The proximate and mineral compositions of the fish were also analyzed. Data were subjected to one-way analysis of variance and Tukey’s test at p < 0.05. Results: The 75% BSF and 50% MW diets exhibited the best growth performance, with significant improvements in WG, FCR, and SGR. Protein digestibility improved significantly, rising from 49.22% (control) to 75.15% in the BSF 75% group and from 50.91% (control) to 75.33% in the MW 50% group. BSF inclusion reduced body fat while increasing protein and ash content, whereas MW showed variable effects on lipid content. Both insect meals enhanced mineral retention, particularly calcium, sodium, phosphorus, and magnesium, with peak values observed in the 75% BSF and 50% MW diets. Conclusion: Replacing fishmeal with BSF and MW larvae meals improves the growth, nutrient digestibility, and mineral composition of L. rohita fingerlings. The BSF at 75% and MW at 50% inclusion levels yielded the best results. Therefore, insect meals are viable, sustainable alternatives to fishmeal in aquafeeds, although careful attention to inclusion levels and chitin content is required to optimize benefits. Keywords: Aqua-feed, Aquaculture, Fishmeal, Insect protein, Sustainability. IntroductionThe population pressure on Earth is increasing day by day. The human population is the main cause of urbanization, and according to projections by the United Nations, the human population is expected to reach 9.3 billion by 2050 (Devic et al., 2018). It is estimated that food requirements will increase by approximately 60% globally by 2050 (Subasinghe et al., 2009). Due to population pressure, most countries in the world are facing problems such as protein and food supply shortages (Naylor et al., 2021). To support this large human population, the maximum production of food is necessary. Aquaculture has become a cornerstone of global food security, offering an essential source of protein and nutrition to an ever-growing human population. According to the Food and Agriculture Organization (2022), fish farming now accounts for approximately 46% of global fish production and 52% of the total seafood consumed by humans. Historically, aquaculture was a marginal activity, but it has grown exponentially, particularly in Asia, becoming a significant economic driver and a sustainable alternative to overfishing (Belghit et al., 2019). The aquaculture industry plays a significant role in the production of aquatic animals, such as fish, mollusks, and crustaceans, and the annual growth rate has been recorded at 5.8% (Limbu and Jumanne, 2014; Fao, 2022). Fish farms in most Asian nations not only provide a source of food but also significantly contribute to the socio-economic well-being of many individuals (Mastoraki et al., 2024). The development of the industry has been prompted by various factors, such as the reduction in the wild fish stock and the need to use efficient production to support the population’s nutritional needs. It is also referred to as the world’s fastest-expanding food production sector (Abdel-Tawwab et al., 2020). Aquatic foods contain high levels of proteins and omega-3 polyunsaturated fatty acids (Barroso et al., 2014; Xiao et al., 2018). Small pelagic fish, e.g., sardines and anchovies, play a major role in fishmeal production, and these species are commonly harvested from marine ecosystems. The result of the practice is a decrease in the number of fish that affects marine food webs and endangers biodiversity (Henry et al., 2015). The need to seek alternative sources of proteins arose due to the increased price of fishmeal to avoid expenditure on feeds and ensure the sustainability of fish farms (Nowak et al., 2016). Industry stakeholders are struggling to come up with alternative sources of proteins that can be substituted in the place of fishmeal in the aquaculture diets to overcome these dilemmas (Sánchez-Muros et al., 2016). It is also thought that the types of alternatives provide the nutrients needed to ensure that the fish grow in the most appropriate manner while minimizing the environmental impact and ethical issues (Caimi et al., 2021). Insects have become a potential protein source because of their high nutrient content, extremely high rate of reproduction, and ability to convert organic waste products into high-quality biomass (Elia et al., 2018). Insects contain a high concentration of essential amino acids, lipids, and micronutrients and can be used in aquafeeds (Renna et al., 2017). The black soldier fly (BSF) (Hermetia illucens) and mealworm (MW) larvae (Tenebrio molitor) are the most promising species (Caimi et al., 2021). BSF larvae contain a protein level of 35–46 percent (Meneguz et al., 2018), a lipid level of 19%–37% (Li et al., 2017), an ash value of 11%–28% and saturated fatty acid level of 58%–72%. Likewise, MWs (Tenebrio molitor) can provide a crude protein (CP) content of 47%–60%. The mean values of fat and ash in MWs were 30.8% and 4.2%, respectively. Crude fiber is 4.19–22.35 g/100 g (Kishawy et al., 2022). Labeo rohita, commonly known as rohu, is one of the key freshwater aquaculture species in South Asia. It is extensively cultivated because of its quick growth, market demand, and consumer preference due to its nutritious quality. Ecologically, rohu plays a critical role in polyculture systems because of its ability to make efficient use of natural food resources in the water column. From an economic standpoint, rohu is a major component of carp aquaculture production in countries such as Pakistan, India, and Bangladesh. Its established use in nutrition, growth performance, and molecular biology research makes it an ideal experimental model for evaluating alternative protein sources, such as insect meal, in aquaculture diets. The inclusion levels (0%, 25%, 50%, 75%, and 100%) of BSF and MW larvae meal were chosen according to previous studies on the gradual replacement of fishmeal with insect meal to predict its effects on growth, digestibility of nutrients, and general health state in fish. These inclusion levels were selected to span a wide array of potential practical uses ranging from low to full replacement of fishmeal, representing different stages of adoption of insect meal in aquaculture. Additionally, these levels enabled a thorough assessment of dose-dependent responses, from a low inclusion (25%) to a high inclusion (100%) that allowed a thorough understanding of the incremental effects of insect meal on fish performance and diet efficiency. Although the possible use of insect protein as a feed ingredient in fish farming has been investigated in various studies, little research has been conducted to investigate its effects on L. rohita. Labeo rohita is used in aquaculture and has acquired economic significance owing to its fast growth rate, tolerance to diverse environments, and high consumer demand within South Asia (Onsongo et al., 2018). This study was undertaken to assess the effect of substituting fish meal with insect meal (BSF and MW larvae) in the diet of L. rohita fingerlings. Specifically, we examined the growth performance, nutrient digestibility, and body and mineral composition of L. rohita. The present results shed light on the prospects of insect protein as a sustainable and cost-effective alternative to conventional fish meal in carp aquaculture. Materials and MethodsExperimental procedureLabeo rohita fingerlings were obtained from Fish Nursery Unit, Head Balloki, Punjab, Pakistan (initial mean weight, 5.56 ± 0.04 g). Fingerlings were carefully transported in water-filled aerated bags. Before the start of the experiment, the fish were immersed in NaCl (5 g/l) solution for a few minutes to remove any ectoparasites. Fingerlings were allowed to acclimatize in glass aquariums under laboratory conditions for 2 weeks. Labeo rohita fingerlings (n=10) were stocked in each aquarium. During the acclimatization period, all fish received the same food ration (3%–5% BW/d) at one feeding daily (Liland et al., 2021). The Institutional Animal Ethics Committee, University of Lahore, Pakistan, approved the experimental protocol. Diet preparationThe experimental diet for the study was prepared in several stages, starting with the basal control diet and culminating in the formulation of the experimental diet containing BSF larvae and MWs as test ingredients. The BSF and MW larvae meals used in this study were purchased in dried powder form from a local supplier (J & J Bird Supplies, Lahore, Pakistan; https://jandjbirdsupplies.com). A basal control diet was prepared to ensure consistency across all treatments. The control diet was developed using standard ingredients including fishmeal, soybean meal, wheat flour, sunflower meal, canola meal, and corn gluten meal, along with essential vitamins and minerals. These ingredients were weighed according to the required percentages and thoroughly mixed using a commercial mixer to ensure that all components were thoroughly combined to form a uniform base mixture. For the experimental diet, the BSF larvae and MWs were ground to a fine powder using a grinder to facilitate better integration into the diet. Powdered BSF larvae and MW meal were then gradually added to the basal diet mixture at varying inclusion levels, typically ranging from 5% to 20%, depending on the experimental treatment. After heating, a small amount of water was added to the mixture to facilitate dough formation. Water was gradually added while the ingredients were continuously mixed to prevent clumping. The consistency of the dough was adjusted to ensure that it was firm but pliable enough to shape into pellets. The dough was then processed through a pelletizer, where it was extruded into small pellets. The pellets were formed and collected for the next stage of drying. After freeze-drying, the pellets were stored in airtight containers to prevent moisture absorption. The containers were kept in a cool, dry place to maintain the diet’s quality and stability. Tables 1 and 2 show the composition of the experimental diets. Table 1. Composition of diets based on varying levels of BSF.
Table 2. Composition of varying levels of MW-based diet (%)/100 g.
Growth trialTwo trials were carried out separately, with 15 aquaria used for each trial. Both trials were simultaneously conducted under the same environmental conditions in the same facility to make them comparable. All aquaria were kept under the same water quality management regime, photoperiod, and feeding schedule to reduce possible tank effects. After the acclimatization period, 10 uniform-sized fish (initial mean weight, 5.56 ± 0.04 g) in good health condition were stocked in each of the 15 aquaria. Each aquarium had a capacity of approximately 311 l with dimensions of 122 × 45.7 × 55.8 cm. Three replicate tanks were used to perform the experiment (n=3 tanks, 10 fish per tank), and each dietary treatment was carried out in three tanks. In the first trial, five diets were prepared by replacing fish meal with BSF larvae meal as follows: 0%, 25%, 50%, 75%, and 100%. In the second trial, five diets were prepared by replacing fish meal with MW larvae meal. Each experiment trial was performed in triplicates for 60 days under a completely randomized design (Fig. 1). The fish were fed two equal portions per day at 0800 and 1,400 hours for 60 days. The experimental trial was conducted at ambient temperature and subjected to a natural photoperiod of approximately 12 hours light/12 hours dark period. Approximately 50% of the water in each aquarium was changed daily with freshwater dechlorinated to provide optimum environmental conditions. The water quality parameters were checked every 3 days according to the standard protocol. At the end of the experiment, all fish from the aquariums were counted and collected for further processing.
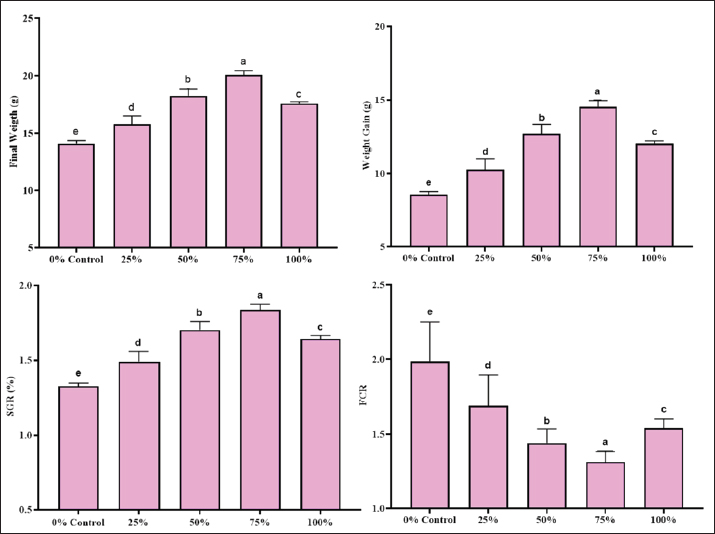
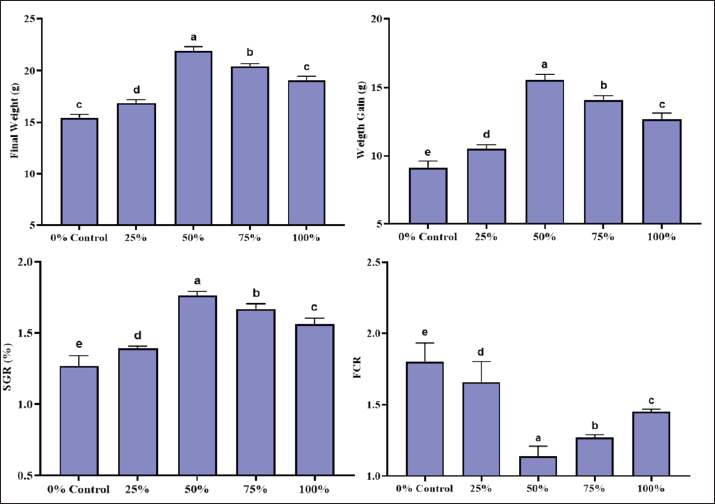
Fig. 1. The aquarium setup used for the experimental trial. Digestibility trial and apparent digestibility coefficients (ADC) determinationsThe digestibility trial was conducted just after the 60-day feeding trial with the same groups of fish. This timing was selected to allow sufficient time for the fish’s intestinal physiology and microbial community to adapt to the experimental diets. Considering the influence of dietary transition on nutrient utilization, conducting the digestibility trial at this stage minimized dietary transition variability and will give a more accurate estimate of nutrient use from test diets. A total of (n=20) fish from each trial with similar body weight were selected and transferred to two aquariums (122 × 45.7 × 55.8 cm, 284 l water) and fed the experimental diets based on the same ration level given in the growth trial. The digestibility trial was continued for 21 days. The daily provided feed was recorded, and uneaten feed was collected 20 minutes after feeding to avoid contamination of feces with uneaten feed. Only the uneaten feed was siphoned, collected on pre-weighed filter paper, dried, and reweighed to calculate the actual feed intake. This procedure ensured that only the consumed feed was used in the growth and digestibility calculations. Feces were gently siphoned from the aquarium onto a 150-mm screen, rinsed with distilled water to remove residual salts, and stored at −20°C for further analysis (Ward et al., 2003). Feces were sampled by gentle siphoning from each tank after a 24-hour feeding period to reduce contamination with uneaten feed. Siphoning may cause leaching of soluble nutrients, such as amino acids, into the water. To reduce this concern, feces were collected as soon as they were defecated, and the tanks were closely monitored during collection sessions. The water quality in each aquarium was maintained by replacing the water once every 2 days before the first feeding. The water quality parameters, including temperature, pH, dissolved oxygen, and ammonia-N, were checked on a daily basis. Fecal samples were ground with a mechanical mortar and pestle and sieved using a 1 mm screen for further analysis. The energy of the sample was determined using an adiabatic bomb calorimeter (Philipson Micro-bomb calorimeter: Gentry Instruments, Aiken, SC, USA) following the standard procedures (AOAC, 2019). As chitin is an indigestible component of insect meal, it was not included in the protein, lipid, and energy digestibility calculations. After removing chitin, the nutrient dietary composition of the diets was used to calculate the digestibility coefficients for protein, lipid, and energy. The ADCs of protein, lipid, and gross energy were calculated following the method of Luo et al. (2006). Body compositionFive fish of the same size were sampled from each experimental aquarium. Sampling was performed on the last day of the experiment after 24 hours of feed withdrawal to reduce the interference of gut content in subsequent analyses. Fishes were oven-dried at 70°C and ground to a powder, which was then analyzed for proximate composition. The proximate analysis of fishes in terms of moisture, total ash, crude fat, and CP was performed to evaluate the nutritive value of fishes according to AOAC (2019). Moisture was determined by oven drying at 105°C for 18 hours, and CP was determined by micro-Kjeldhal analysis. Crude fat content was determined according to a standard protocol (Luo et al., 2006). Mineral compositionFish samples (10 g) were ashed at 550°C for 6 hours in a muffle furnace. The ashed sample was diluted in 5 ml of 10% HCl. This was filtered and made up to the mark in 50 analysis flask, and the filtrate was used for the analysis. The mineral composition of the fish was analyzed according to the methods of AOAC (2019). Statistical analysisThe obtained data were subjected to one-way ANOVA, and Tukey’s honesty significant difference test was applied to compare differences among means. The level of significance was chosen at p < 0.05 using CoStat (version 6.303) (Steel et al., 1996). Ethical approvalNot needed for this study. ResultsGrowth performanceThe growth performance of L. rohita was tested in two diet groups that contained different proportions of BSF and MW larvae meal replacing fishmeal. The findings indicated that the most significant growth performance was in the 75% BSF and 50% MW diet groups. Growth performance was measured using the following parameters: weight gain (WG), feed conversion ratio (FCR), and specific growth rate (SGR). Initially, L. rohita fingerlings had an average initial weight of 5.56 ± 0.04 g across all treatment groups. The 75% BSF-based diet led to the highest WG, SGR, and FCR of 14.53 ± 0.43 g, 1.837 ± 0.038, and 1.313 ± 0.071, respectively (Fig. 2). Similarly, the data showed that the 50% MW-based diet group had the best growth performance. The maximum WG and better SGR and FCR were recorded in the 50% MW based diet group as 15.56 ± 0.41 g, 1.763 ± 0.029, and 1.140 ± 0.070, respectively (Fig. 3).
Fig. 2. Growth performance of L. rohita fed on different levels of BSF-based diet.
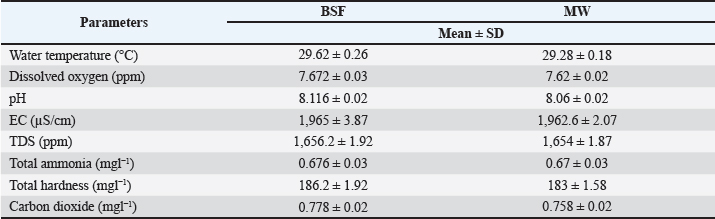
Fig. 3. Growth performance of L. rohita fed on different MW-based diet levels. Parameters of water qualityWater quality is a critical parameter in aquaculture as it has an impact on fish health and performance. Water quality parameters, such as temperature, dissolved oxygen, pH, electric conductivity (EC), total dissolved solids (TDSs), total ammonia, total hardness, and carbon dioxide, were measured on a fortnightly basis throughout the experiment. The results reveal that the water quality parameters fell within the optimal range of growth of L. rohita in the three treatment groups, and no significant differences were observed between the BSF and MW diets. The mean water temperature was 29.62°C ± 0.26°C and 29.28°C ± 0.18°C in the BSF and MW diet groups, respectively. These parameters were within the acceptable range for L. rohita growth. The dissolved oxygen levels were slightly higher in the BSF group (7.672 ± 0.03 ppm) than in the MW group (7.62 ± 0.02 ppm). The pH levels were nearly the same in both diet groups, as 8.116 ± 0.02 and 8.06 ± 0.02 in the BSF and MW diet groups, respectively. The electrical conductivity values were nearly the same in both the BSF and MW groups at 1965 ± 3.87 and 1962.6 ± 2.07 µS/cm, respectively. The TDS values were 1,656.2 ± 1.92 and 1,654 ± 1.87 ppm in the BSF and MW diet groups, respectively. The recorded total ammonia and hardness values were within the optimum range, ensuring that no toxic effect of ammonia and water hardness was applied to fish health (Table 3). Table 3. Water quality parameters during the experimental period.
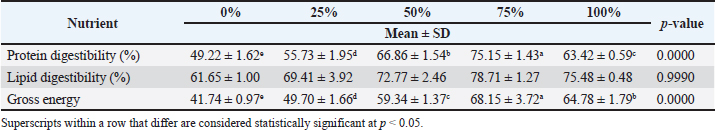
Nutrient digestibilityNutrient digestibility was measured in terms of protein, fat, and energy. Significant differences in protein and gross energy digestibility were observed among the different diet groups. The results indicated a marked improvement in nutrient digestibility in the BSF and MW meal groups, especially in the BSF 75% and MW 50% groups (Tables 4 and 5). Table 4. Obvious nutrient digestibility (protein, lipid, and gross energy) of L. rohita fingerlings fed with diets containing varying amounts of BSF meal.
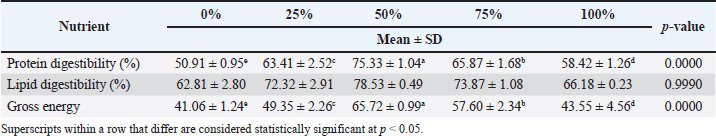
Table 5. Obvious nutrient digestibility (protein, lipid, and gross energy) of L. rohita fingerlings fed on diets with varying levels of MW meal.
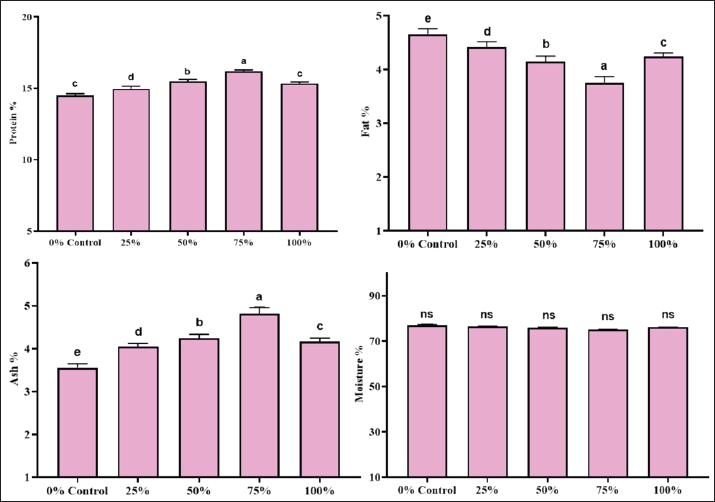
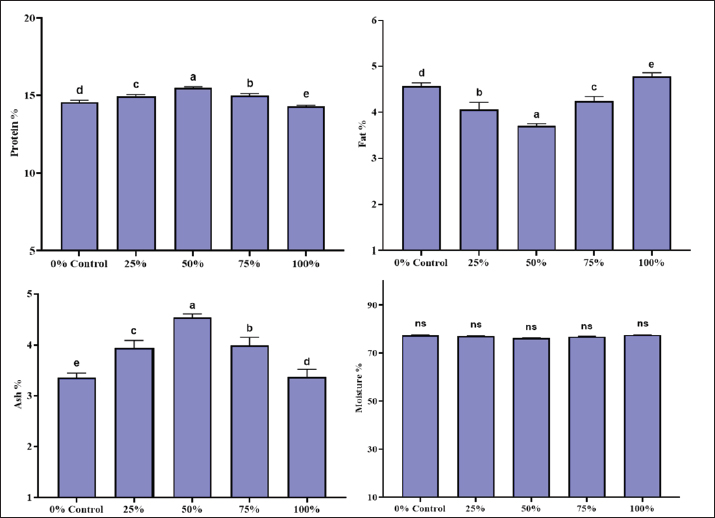
Nutrient digestibility in the BSF groupProtein digestibility significantly increased with the inclusion of BSF meal. The control group (0% BSF) had the lowest protein digestibility at 49.22% ± 1.62%, whereas the BSF 75% group exhibited the highest protein digestibility at 75.15% ± 1.43%. The BSF 50% group also showed a notable improvement in protein digestibility (66.86% ± 1.54%), which was significantly higher than that of the control group. The p-value for protein digestibility was 0.0000, indicating a highly significant difference across the groups. The control group had a lipid digestibility of 61.65% ± 1.00%, and the BSF 75% group had the highest digestibility at 78.71% ± 1.27%. However, the p-value for lipid digestibility was 0.9990, suggesting that these differences were not statistically significant. Gross energy showed a clear increasing trend with BSF inclusion, with the BSF 75% group showing the highest digestibility (68.15% ± 3.72%), followed by the BSF 50% group (59.34% ± 1.37%). The control group exhibited the lowest gross energy digestibility at 41.74% ± 0.97%). The p-value for gross energy digestibility was 0.0000, indicating a statistically significant difference between the groups. Nutrient digestibility in the MW groupThe protein digestibility increased significantly with the inclusion of the MW meal. The control group (0% MW) had the lowest protein digestibility at 50.91% ± 0.95%, whereas the MW 50% group exhibited the highest protein digestibility at 75.33% ± 1.04%. The MW 25% and MW 75% groups showed intermediate values of 63.41% ± 2.52% and 65.87% ± 1.68%, respectively. The p-value for protein digestibility was 0.0000, indicating a statistically significant difference across the groups. Lipid digestibility also improved with increasing MW meal levels, but the differences were not statistically significant. The control group had a lipid digestibility of 62.81% ± 2.80%, and the MW 50% group had the highest value at 78.53% ± 0.49%. The p-value for lipid digestibility was 0.9990, suggesting no significant differences between the groups. Gross energy showed a clear increasing trend with MW inclusion, with the 50% MW group showing the highest digestibility (65.72% ± 0.99%), followed by the 25% MW group (49.35% ± 2.26%). The control group showed the lowest gross energy digestibility at 41.06% ± 1.24%. The p-value for gross energy digestibility was 0.0000, indicating a statistically significant difference between the groups. Body compositionThe body composition analysis included the measurement of proximate composition, such as moisture, protein, fat, and ash contents. The body composition of L. rohita was significantly influenced by the inclusion of BSF and MW larvae meal (Figs. 4 and 5).
Fig. 4. Body composition of L. rohita fed with different BSF-based diet levels.
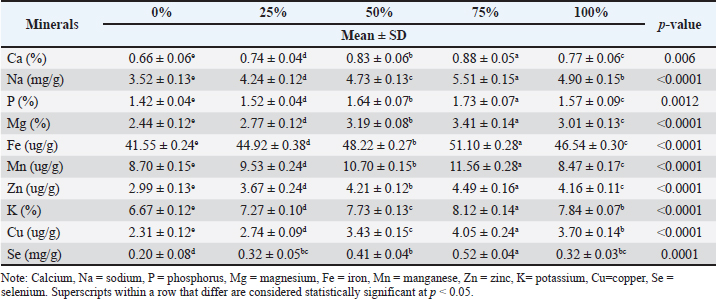
Fig. 5. Body composition of L. rohita fed on different MW-based diet levels. Protein contentThe protein content of L. rohita showed slight variations across the different diet groups. In the BSF diet groups, protein content increased with the inclusion of BSF meal, from 14.51% ± 0.13% in the control group (0% BSF) to 16.19% ± 0.10% in the BSF 75% group. The BSF 100% group had a protein content of 15.34% ± 0.11%, which was slightly lower than that of the BSF 75% group. In contrast, the protein content remained relatively stable in the MW diet groups, with a slight increase from 14.56% ± 0.14% in the control group (0% MW) to 15.49% ± 0.06% in the MW 50% group. However, the MW 100% group showed a decrease in protein content to 14.30% ± 0.06%. Lipid contentThe fat content in L. rohita varied across the different diet groups. In the BSF diet groups, the fat content decreased with increasing levels of BSF meal, from 4.66% ± 0.10% in the control group (0% BSF) to the lowest value of 3.75% ± 0.12% in the 75% BSF group. BSF 25% and 100% groups had intermediate fat contents of 4.42% ± 0.10% and 4.24% ± 0.07%, respectively. In contrast, the fat content in the MW diet groups exhibited a more fluctuating pattern. The control group (0% MW) had a fat content of 4.56% ± 0.08%, which decreased to 3.70% ± 0.05% in the 50% MW group. However, the MW 75% group showed an increase in fat content to 4.24% ± 0.10%, and the MW 100% group had the highest fat content at 4.78% ± 0.08%. These results suggest that the BSF diet tends to reduce fat content as the inclusion level increases, whereas the MW diet showed more variation with no clear trend. Ash contentThe ash content in the BSF diet group tended to increase with the BSF meal inclusion. The control group (0% BSF) had the lowest ash content (3.56% ± 0.09%), whereas the BSF 75% group had the highest ash content (4.82% ± 0.14%). The BSF 25%, 50%, and 100% groups showed intermediate values of 4.05% ±0.08%, 4.25% ± 0.09%, and 4.17% ± 0.08%, respectively. In contrast, the MW diet groups lacked consistency in terms of ash content. The lowest ash content (3.36% ± 0.09%) was recorded in the control group (0% MW) and highest was (4.54% ± 0.07%) in the MW 50% group. However, the MW 25% and 75% 100% groups showed intermediate results as 3.94% ± 0.15%, 3.99% ± 0.16%, and 3.37% ± 0.15%, respectively. These results indicated that the BSF meal was more effective in increasing ash content, whereas the MW diet showed more variation between the diet groups. Moisture contentThere was a slight difference in the moisture content of L. rohita across the different diet groups, but there were no significant differences between the BSF and MW diet groups. The moisture content was 77.27% ± 0.11%, 76.54% ± 0.21%, 75.24% ± 0.070%, and 76.13% ± 0.21% in the 0%, 25%, 50%, 75%, and 100% BSF diet groups, respectively. Similarly, the moisture content showed low variation in the MW diet groups, ranging from 77.05% ± 0.18% (MW 25%) to 76.79% ± 0.18% in the MW 75% group. Overall, these results indicate that neither the BSF nor the MW meal inclusion significantly affected the moisture content of L. rohita, with values remaining relatively consistent across all diet groups. Mineral compositionLabeo rohita fingerlings were evaluated based on the mineral content of calcium (Ca), sodium (Na), phosphorus (P), magnesium (Mg), iron (Fe), manganese (Mn), zinc (Zn), potassium (K), copper (Cu), and selenium (Se) after feeding on MW and BSF larvae diets of different concentrations. The groups differed significantly, particularly regarding the levels of calcium, sodium, phosphorus, and magnesium, which increased with increased BSF meal inclusion (Tables 6 and 7). Table 6. Mineral composition of L. rohita fingerling fed with varying levels of BSF diet.
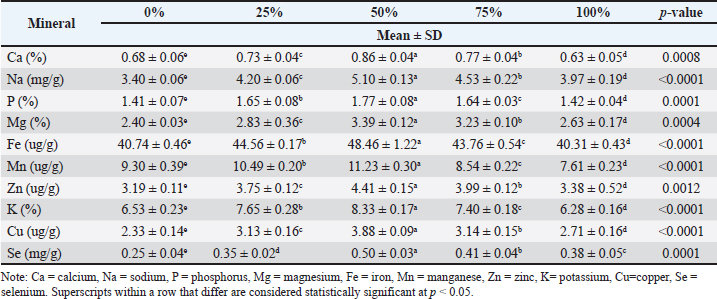
Table 7. Mineral composition of L. rohita fingerling fed with varying levels of MW diet.
Calcium (Ca)The calcium content increased significantly with the inclusion of BSF meal. The control group (0% BSF) had the lowest calcium content at 0.66% ± 0.06%, whereas the BSF 75% group exhibited the highest value at 0.88% ± 0.05%. The 25%, 50%, and 100% BSF groups had intermediate values of 0.74% ± 0.04%, 0.83% ± 0.06%, and 0.77% ± 0.06%, respectively. The p-value for calcium was 0.006, indicating a significant difference between the groups. The calcium content significantly increased with the inclusion of the MW meal. The control group (0% MW) had the lowest calcium content (0.68% ± 0.06%) and the 50% MW group had the highest calcium content (0.86% ± 0.04%), whereas the 25%, 75%, and 100% MW groups showed intermediate values of 0.73% ± 0.04%, 0.77% ± 0.04%, and 0.63% ± 0.05%, respectively. The p-value for calcium was 0.0008, indicating a significant difference. Sodium (Na)The sodium level significantly increased with higher BSF meal inclusion (p < 0.0001). The control group had the lowest sodium level (3.52 ± 0.13mg/g), and the highest was recorded in the BSF 75% group (5.51 ± 0.15mg/g). The 25%, 50%, and 100% BSF groups had intermediate levels of 4.24 ± 0.12, 4.73 ± 0.13, and 4.90 ± 0.15 mg/g, respectively. Sodium levels also significantly increased with higher MW meal inclusion (p < 0.0001). The control group had 3.40 ± 0.06 mg/g of sodium, which increased to 5.10 ± 0.13 mg/g in the MW 50% group. The 25%, 75%, and 100% MW groups showed intermediate sodium levels of 4.20 ± 0.06, 4.53 ± 0.22, and 3.97 ± 0.19 mg/g, respectively. The p-value for sodium was (<0.0001), indicating a highly significant difference. Phosphorus (P)Phosphorus content increased significantly from 1.42% ± 0.04% in the control group to 1.73% ± 0.07% in the 75% BSF group (p < 0.0012). Intermediate values of 1.52%, 0.04%, 50%, and 100% BSF groups at 1.52% ± 0.04%, 1.64% ± 0.07%, and 1.57% ± 0.09%, respectively. The phosphorus content increased significantly from 1.41% ± 0.07% in the control group to 1.77% ± 0.08% in the MW 50% group (p < 0.0001). The 25%, 75%, and 100% MW groups had 1.65% ± 0.08%, 1.64% ± 0.03%, and 1.42% ± 0.04%, respectively. Magnesium (Mg)The magnesium content increased significantly with the inclusion of BSF meal (p < 0.0001). The control group had the lowest magnesium level (2.44% ± 0.12%), whereas the BSF 75% group had the highest (3.41% ± 0.14%). The 25%, 50%, and 100% BSF groups showed intermediate values of 2.77% ± 0.12%, 3.19% ± 0.08%, and 3.01% ± 0.13%, respectively. Magnesium content increased significantly with the inclusion of MW meal, from 2.40% ± 0.03% in the control group to 3.39% ± 0.12% in the MW 50% group. The 25%, 75%, and 100% MW groups had intermediate values of 2.83% ± 0.36%, 3.23% ± 0.10%, and 2.63% ± 0.17%, respectively. The p-value for magnesium was 0.0004, indicating a significant difference. DiscussionFishmeal is a high-protein feed ingredient typically made of wild fish, and commercial fish feeds heavily rely on fishmeal. Harvesting fishmeal poses a major sustainability issue, especially to marine life. Excessive exploitation of small pelagic fish for use in fishmeal production can cause an imbalance in the food chain, with ripple effects on other marine organisms. The importance of finding alternative and sustainable sources of feed ingredients that will reduce the dependence on fishmeal is increasingly being realized (Dai et al., 2023; Abdel-Tawwab et al., 2025). This involves researching plant-based proteins and insect meals that are environmentally friendly (Haarhaus et al., 2022). The findings of the present study have elucidated the role of BSF and MW larvae as potential substitutes for fishmeal in the diet of L. rohita. The results are consistent with those of previous studies in which positive changes in digestibility and growth performance of fish fed on insect-based diets were observed. The incorporation of 75% BSF and 50% MW as dietary supplements in the diet of L. rohita was found to improve growth performance and feed efficiency, thereby registering them as viable alternatives to the conventional fishmeal-based diets. BSF larvae meal diet has been tested in a variety of fish species such as European seabass by Abdel-Latif et al. (2021) rainbow trout (Oncorhynchus mykiss) (Renna et al., 2017; Elia et al., 2018), juvenile rainbow trout (Oncorhynchus mykiss) by St-Hilaire et al. (2007), Atlantic salmon (Salmo salar) by (Belghit et al., 2018), Jian carp (Cyprinus carpio var. Jian) (Li et al., 2017; Zhou et al., 2018), European seabass (Dicentrarchus labrax) (Magalhães et al., 2017), Nile tilapia (Oreochromis niloticus) by Kishawy et al. (2022) postsmolt Atlantic salmon (Lock et al., 2016; Belghit et al., 2019), and fingerlings of Nile tilapia (Devic et al., 2018) and European seabass (Dicentrarchus labrax) (Abdel-Tawwab et al., 2020). Even with these findings, the substitution of FM with BSF larvae meal decreased the growth of juvenile turbot (Psetta maxima) (Kroeckel et al., 2012) and subadult channel catfish (Ictalurus punctatus), where 100% BSF larvae lowered feed consumption (Bondari and Sheppard, 1987). The present findings demonstrated that the partial replacement of fishmeal with BSF and MW larvae meals significantly improved the growth performance, nutrient digestibility, and mineral composition of L. rohita. Similar results were reported by Belghit et al. (2019) and Abdel-Tawwab et al. (2020), who found that BSF inclusion enhanced the growth rate and feed conversion efficiency in Atlantic salmon and European sea bass, respectively. The observed increase in lipid digestibility with the addition of higher levels of insect meal inclusion suggests that L. rohita may be more adapted to utilize the lipids in insect-based diets due to the potential for different fatty acid profiles and lipid metabolism pathways associated with these meals. This could be correlated with the higher proportion of unsaturated fatty acids found in BSF and MW larvae, which could be responsible for the improved digestibility by increasing the emulsification and absorption process in the gastrointestinal tract of the fish. In addition, the high fat content of insect-based diets may promote better lipid utilization efficiency in L. rohita, which is characterized by its ability to efficiently metabolize lipid-rich diets in the juvenile stage. These results support the idea that insect meals, which are high in essential fatty acids, could be of a more digestible and beneficial lipid profile to aquaculture applications, potentially reducing the need for fish oil and other conventional lipid sources. In terms of body composition, the protein content of the fish improved when the fish were fed on higher levels of BSF inclusion. These same tendencies were also observed in the MW diet groups, although they were lower. These findings align with those of Hameed et al. (2022), who also found that fish maintained an increase in protein retention when insect-based diets were used. However, the fat content in L. rohita decreased with BSF addition, although there was variability in the MW diet. Such differences in lipid content indicate that insect meal type (BSF or MW) can affect fat deposition in fish, with the BSF meal leading to fewer fat deposits. The results of the mineral composition showed that there was an increased level of calcium, sodium, phosphorus, and magnesium in the BSF and MW diet groups. These minerals are critical to fish growth, bone development, and general health. The higher levels of calcium and magnesium, especially in the 75% BSF group, are encouraging and in line with the study by Sharifinia et al. (2025), who reported higher mineral retention in shrimp fed with insect-based diets. This confirms that insect meals not only supply essential nutrients but also contribute to better mineral absorption, possibly due to the exoskeleton’s bioavailability of trace elements. Mastoraki et al. (2020) also mentioned that diets high in fat, especially from BSF, can influence liver size and function. Shin and Lee (2021) reported similar findings in shrimp, where higher BSF inclusion led to changes in lipid metabolism and liver function, further supporting our results. Our study also contributes to the growing body of literature on the environmental benefits of using insect meal in aquafeeds. Insect farming is known for its low ecological footprint, as insects can be raised on organic waste, using far fewer resources such as land and water compared to traditional feed ingredients such as soy and fishmeal (Wu et al., 2022). Magalhães et al. (2017) emphasized that insect meal production significantly reduces the environmental impact of aquaculture by promoting waste recycling and offering an eco-friendly protein alternative. The implications of our findings are far-reaching. Insect meals, particularly from BSF, offer a sustainable, nutritious alternative to fishmeal, with potential economic benefits for the aquaculture industry. As insect farming continues to expand, the availability of insect protein will increase, further driving down costs while improving aquafeed production sustainability. This is especially important given the increasing demand for aquaculture products and the associated environmental concerns. However, several areas warrant further investigation. First, the long-term effects of insect meal inclusion on fish health and reproductive performance remain unclear, and these aspects should be addressed in future studies. In addition, species-specific responses to different insect meals should be explored, as the digestibility of chitin and other components may vary across species. Future research should also investigate the optimal insect meal inclusion levels to avoid potential negative effects, particularly on liver health and growth performance at high inclusion levels. Yu et al. (2020) reported that replacing 25% of fishmeal with BSF larval meal improved growth performance and feed intake in golden pompano (Trachinotus ovatus). Similarly, Sharifinia et al. (2023) found that incorporating MW larval meal into the diet of Pacific white shrimp (Litopenaeus vannamei) enhanced both growth performance and immune system responses. These studies align with our results, which indicate that BSF larval meal, particularly at a 75% inclusion level, significantly improved nutrient digestibility and growth performance in L. rohita. The enhanced digestibility observed in our study is consistent with the findings of Weththasinghe et al. (2022), who noted that full-fat BSF larval meal could replace up to 12.5% of dietary fishmeal without compromising nutrient digestibility. Additionally, Chen et al. (2025) reported that yellow MW larvae meal improved growth performance and hepatic health in juvenile LB (Micropterus salmoides). Collectively, these findings underscore the potential of insect larvae, especially BSF and MW, as sustainable and nutritionally efficient alternatives to traditional fishmeal in aquaculture diets. Their use not only improves growth and nutrient digestibility but also contributes to the sustainability of aquaculture practices by lowering the use of fishmeal sourced from wild fish stocks. Although this study revealed the nutritional potential of insect meal in L. rohita, an economic study of feed formulation has not been conducted. In Pakistan, the market prices of BSF and MW meals fluctuate, and cost structures will alter as production increases. A cost-benefit analysis and biological examination should be incorporated in future research to justify the mass implementation of aquaculture. This experiment lasted more than 60 days, which is a normal period for assessing the growth and nutrient consumption of L. rohita fingerlings. Nevertheless, more extended research is required to investigate the possible impacts of insect meal addition on broodstock performance, reproductive indices, and multigenerational performance. These studies would be significant for the sustainable use of insect protein in carp farming. ConclusionThe findings of the present study revealed that the inclusion of BSF and MW meal diets for L. rohita improved growth performance, nutrient digestibility, and body and mineral composition. Further research is required to evaluate the long-term effects on fish health, immune system, and reproduction. In addition, the level of inclusion and chitin content in insect-based diets should be carefully considered to ensure maximum advantages and minimum adverse effects on growth and digestibility. Conflict of interestThe authors declare no conflicts of interest. FundingThis study received no specific grant. Authors' contributionsNida Ismat: Methodology, Investigation, Writing-Original Draft, Sikandar Hayat: Conceptualization, Supervision, Formal analysis, Majid Hussain: Writing–Review & Editing, Data Curation. Data availabilityData supporting the findings of this study are not openly available due to sensitivity reasons. However, they are available upon reasonable request from the corresponding author. ReferencesAbdel-Latif, H.M.R., Abdel-Tawwab, M., Khalil, R.H., Metwally, A.A., Shakweer, M.S., Ghetas, H.A. and Khallaf, M.A. 2021. Black soldier fly (Hermetia illucens) larvae meal in diets of European seabass: effects on antioxidative capacity, non-specific immunity, transcriptomic responses, and resistance to the challenge with Vibrio alginolyticus. Fish Shellfish Immunol. 111, 111–118. Abdel-Tawwab, M., Khalil, R.H., Metwally, A.A., Shakweer, M.S., Khallaf, M.A. and Abdel-Latif, H.M.R. 2020. Effects of black soldier fly (Hermetia illucens L.) larvae meal on growth performance, organs-somatic indices, body composition, and hemato-biochemical variables of European sea bass, Dicentrarchus labrax. Aquaculture 522, 735–136. Abdel-Tawwab, M., Khalil, R.H., Selema, T.A.A., Abdelsalam, M., Abdelhakim, T.M., Sabry, E.A. and Saad, A.H. 2025. Evaluating the inclusion of Clostridium autoethanogenum protein instead of fishmeal protein in diets for European seabass (Dicentrarchus labrax): growth performance, digestive enzymes, health status, and tissues investigations. Animals 324, 116318. AOAC. 2019. Official methods of analysis of the Association of Official Analytical Chemists: official Methods of Analysis of AOAC International. Washington, DC: AOAC. Barroso, F.G., De Haro, C., Sánchez-Muros, M.J., Venegas, E., Martínez-Sánchez, A. and Pérez-Bañón, C. 2014. The potential of various insect species for use as food for fish. Aquaculture 422–423(423), 193–201. Belghit, I., Liland, N.S., Gjesdal, P., Biancarosa, I., Menchetti, E., Li, Y., Waagbø, R., Krogdahl, A. and Lock, E.J. 2019. Black soldier fly larvae meal can replace fish meal in diets of sea-water phase Atlantic salmon (Salmo salar). Aquaculture 503, 609–619. Belghit, I., Liland, N.S., Waagbø, R., Biancarosa, I., Pelusio, N., Li, Y., Krogdahl, A. and Lock, E.J. 2018. Potential of insect-based diets for Atlantic salmon (Salmo salar). Aquaculture 491, 72–81. Bondari, K. and Sheppard, D.C. 1987. Soldier fly, Hermetia illucens L., larvae as feed for channel catfish, Ictalurus punctatus (Rafinesque), and blue tilapia, Oreochromis aureus (Steindachner). Aquac. Res. 18(3), 209–220. Caimi, C., Biasato, I., Chemello, G., Oddon, S.B., Lussiana, C., Malfatto, V.M., Capucchio, M.T., Colombino, E., Schiavone, A., Gai, F., Trocino, A., Brugiapaglia, A., Renna, M. and Gasco, L. 2021. Dietary inclusion of a partially defatted black soldier fly (Hermetia illucens) larva meal in low fishmeal-based diets for rainbow trout (Oncorhynchus mykiss). J. Anim. Sci. Biotechnol. 12, 50. Chen, Y., Li, W., Zhong, M., Ma, J., Chen, B., Cao, J., Loh, J.Y. and Huang, H. 2025. Black soldier fly larvae meal as a sustainable fishmeal substitute for juvenile hybrid grouper: impacts on growth, immunity, and gut health. Fishes 10(7), 344. Dai, J., Chen, T., Guo, X., Dai, Z., He, Z. and Hu, Y. 2023. Evaluation of fish meal replacement by Clostridium autoethanogenum protein in diets for juvenile red swamp crayfish (Procambarus clarkii). Aquaculture 570, 739–379. Devic, E., Leschen, W., Murray, F. and Little, D.C. 2018. Growth performance, feed utilization and body composition of advanced nursing Nile tilapia (Oreochromis niloticus) fed diets containing black soldier fly (Hermetia illucens) larvae meal. Aquac. Nutr. 24(1), 416–423. Elia, A.C., Capucchio, M.T., Caldaroni, B., Magara, G., Dörr, A.J.M., Biasato, I., Biasibetti, E., Righetti, M., Pastorino, P., Prearo, M., Gai, F., Schiavone, A. and Gasco, L. 2018. Influence of Hermetia illucens meal dietary inclusion on the histological traits, gut mucin composition and the oxidative stress biomarkers in rainbow trout (Oncorhynchus mykiss). Aquaculture 496, 50–57. FAO. 2022. The state of world fisheries and aquaculture. Toward blue transformation. Rome, Italy: FAO; doi:10.4060/cc0461en Haarhaus, M., Cianciolo, G., Barbuto, S., La Manna, G., Gasperoni, L., Tripepi, G., Plebani, M., Fusaro, M. and Magnusson, P. 2022. Alkaline phosphatase: an old friend as treatment target for cardiovascular and mineral bone disorders in chronic kidney disease. Nutrients 14(10), 2124. Hameed, A., Majeed, W., Naveed, M., Ramzan, U., Bordiga, M., Hameed, M., Ur Rehman, S. and Rana, N. 2022. Success of aquaculture industry with new insights of using insects as feed: a review. Fishes 7(6), 395. Henry, M., Gasco, L., Piccolo, G. and Fountoulaki, E. 2015. Review on the use of insects in the diet of farmed fish: past and future. Anim. Feed Sci. Technol. 203, 1–22. Kishawy, A.T.Y., Mohammed, H.A., Zaglool, A.W., Attia, M.S., Hassan, F.A.M., Roushdy, E.M., Ismail, T.A. and Ibrahim, D. 2022. Partial defatted black solider larvae meal as a promising strategy to replace fish meal protein in diet for Nile tilapia (Oreochromis niloticus): performance, expression of protein and fat transporters, and cytokines related genes and economic efficiency. Aquaculture 555, 738195. Kroeckel, S., Harjes, A.G.E., Roth, I., Katz, H., Wuertz, S., Susenbeth, A. and Schulz, C. 2012. When a turbot catches a fly: evaluation of a pre-pupae meal of the black soldier fly (Hermetia illucens) as fish meal substitute growth performance and chitin degradation in juvenile turbot (Psetta maxima). Aquaculture 364, 345–352. Li, S., Ji, H., Zhang, B., Zhou, J. and Yu, H. 2017. Defatted black soldier fly (Hermetia illucens) larvae meal in diets for juvenile Jian carp (Cyprinus carpio var. Jian): growth performance, antioxidant enzyme activities, digestive enzyme activities, intestine and hepatopancreas histological structure. Aquaculture 477, 62–70. Liland, N.S., Araujo, P., Xu, X.X., Lock, E.J., Radhakrishnan, G., Prabhu, A.J.P. and Belghit, I. 2021. A meta-analysis on the nutritional value of insects in aquafeeds. J. Insects Food Feed 7(5), 743–760. Limbu, S.M. and Jumanne, K. 2014. Effect of restricted and re-feeding regime on feeding cost, growth performance, feed utilization and survival rate of mixed sex Nile tilapia Oreochromis niloticus cultured in tanks. Int. J. Fish. Aquat. Stud. 2(1), 118–123. Lock, E.R., Arsiwalla, T. and Waagbø, R. 2016. Insect larvae meal as an alternative source of nutrients in the diet of Atlantic salmon (Salmo salar) post-smolt. Aquac. Nut. 22(6), 1202–1213. Luo, Z., Liu, Y.J., Mai, K.S., Tian, L.X., Tan, X.Y. and Shi, J.F. 2006. Effects of feeding levels on growth performance, feed utilization, body composition, and apparent digestibility coefficients of nutrients for grouper Epinephelus coioides juveniles. J. World. Aquac. Soc. 37(1), 32–40. Magalhães, R., Sánchez-López, A., Leal, R.S., Martínez-Llorens, S., Oliva-Teles, A. and Peres, H. 2017. Black soldier fly (Hermetia illucens) pre-pupae meal as a fish meal replacement in diets for European seabass (Dicentrarchus labrax). Aquaculture 476, 79–85. Mastoraki, M., Paudel, K., Biasato, I., Oddon, S.B., Caimi, C., Gasco, L. and Chatzifotis, S. 2024. Impact of processing technologies on insect meal digestibility in rainbow trout (Oncorhynchus mykiss) and European sea bass (Dicentrarchus labrax). J. Insects Food Feed 11(17), 49–60. Mastoraki, M., Vlahos, N., Patsea, E., Chatzifotis, S., Mente, E. and Antonopoulou, E. 2020. The effect of insect meal as a feed ingredient on survival, growth, and metabolic and antioxidant response of juvenile prawn Palaemon adspersus (Rathke, 1837). Aquac. Res. 51(9), 3551–3562. Meneguz, M., Schiavone, A., Gai, F., Dama, A., Lussiana, C., Renna, M. and Gasco, L. 2018. Effect of rearing substrate on growth performance, waste reduction efficiency and chemical composition of black soldier fly (Hermetia illucens) larvae. J. Sci. Food Agric. 98(15), 5776–5784. Naylor, R.L., Hardy, R.W., Buschmann, A.H., Bush, S.R., Cao, L., Klinger, D.H., Little, D.C., Lubchenco, J., Shumway, S.E. and Troell, M. 2021. A 20-year retrospective review of global aquaculture. Nature 591(7851), 551–563. Nowak, V., Persijn, D., Rittenschober, D. and Charrondiere, U.R. 2016. Review of food composition data for edible insects. Food Chem. 193, 39–46. Onsongo, V.O., Osuga, I.M., Gachuiri, C.K., Wachira, A.M., Miano, D.M., Tanga, C.M., Ekesi, S., Nakimbugwe, D. and Fiaboe, K.K.M. 2018. Insects for income generation through animal feed: effect of dietary replacement of soybean and fish meal with black soldier fly meal on broiler growth and economic performance. J. Econ. Entomol. 111, 1966–1973. Renna, M., Schiavone, A., Gai, F., Dabbou, S., Lussiana, C., Malfatto, V., Prearo, M., Capucchio, M.T., Biasato, I., Biasibetti, E., De Marco, M., Brugiapaglia, A., Zoccarato, I. and Gasco, L. 2017. Evaluation of the suitability of a partially defatted black soldier fly (Hermetia illucens L.) larvae meal as ingredient for rainbow trout (Oncorhynchus mykiss Walbaum) diets. J. Anim. Sci. Biotechnol. 8(1), 57. Sánchez-Muros, M., De Haro, C., Sanz, A., Trenzado, C.E., Villareces, S. and Barroso, F.G. 2016. Nutritional evaluation of Tenebrio molitor meal as fishmeal substitute for tilapia (Oreochromis niloticus) diet. Aqua. Nut. 22(5), 943–955. Sharifinia, M., Bahmanbeigloo, Z.A., Keshavarzifard, M., Khanjani, M.H., Daliri, M., Koochaknejad, E., and Jasour, M.S. 2023. Fishmeal replacement by mealworm (Tenebrio molitor) in diet of farmed Pacific white shrimp (Litopenaeus vannamei): effects on growth performance, serum biochemistry, and immune response. Aquat. Living Resour. 36, 19; doi: https://doi.org/10.1051/alr/2023013 Sharifinia, M., Dashtiannasab, A., Mobaraki, S. and Pazir, M.K. 2025. Effects of dietary inclusion of mealworm (Tenebrio molitor) on the fatty acid compositions of Pacific white shrimp (Litopenaeus vannamei). Iran. J. Fish. Sci. 24(4), 983–999. Shin, J. and Lee, K.J. 2021. Digestibility of insect meals for Pacific white shrimp (Litopenaeus vannamei) and their performance for growth, feed utilization and immune responses. PLos One. 16(11), 260305. Steel, R.G.D., Torrie, J.H. and Dinkey, D.A. 1996. Principles and procedures of statistics, St-Hilaire, S., Cranfill, K., McGuire, M.A., Mosley, E.E., Tomberlin, J.K., Newton, L., Sealey, W., Sheppard, C. and Irving, S. 2007. Fish offal recycling by the black soldier fly produces a foodstuff high in omega-3 fatty acids. J. World Aqua. Soc. 38, 309–313. Subasinghe, R., Soto, D. and Jia, J. 2009. Global aquaculture and its role in sustainable development. Rev. Aqua. 1(1), 2–9. Ward, L.R., Carter, C.G., Crear, B.J. and Smith, D.M. 2003. Optimal dietary protein level for juvenile southern rock lobster, Jasus edwardsii, at two lipid levels. Aquaculture 217(1-4), 483–500. Weththasinghe, P., Rocha, S.D.C., Øyås, O., Lagos, L., Hansen, J.Ø., Mydland, L.T. and Øverland, M. 2022. Modulation of Atlantic salmon (Salmo salar) gut microbiota composition and predicted metabolic capacity by feeding diets with processed black soldier fly (Hermetia illucens) larvae meals and fractions. Anim. Microbiome. 4(1), 9. Wu, Y., Tian, S., Yuan, J., Zhang, Z., Zhou, H., Gao, W., Zhang, W. and Mai, K. 2022. Effects of Clostridium autoethanogenum protein as substitute for dietary fishmeal on the growth, feed utilization, intestinal health and muscle quality of large yellow croaker Larimichthys crocea. Aquaculture 561, 738591. Xiao, X., Jin, P., Zheng, L., Cai, M., Yu, Z., Yu, J. and Zhang, J. 2018. Effects of black soldier fly (Hermetia illucens) larvae meal protein as a fishmeal replacement on the growth and immune index of yellow catfish (Pelteobagrus fulvidraco). Aqua. Res. 49(4), 1569–1577. Yu, Y., Wang, Q., Huang, Z., Ding, L. and Xu, Z. 2020. Immunoglobulins, mucosal immunity and vaccination in teleost fish. Front. Immunol. 11, 567941. Zhou, J.S., Liu, S.S., Ji, H. and Yu, H.B. 2018. Effect of replacing dietary fish meal with black soldier fly larvae meal on growth and fatty acid composition of Jian carp (Cyprinus carpio var. Jian). Aqua. Nut. 24(1), 424–433. | ||

| How to Cite this Article |
| Pubmed Style Ismat N, Hayat S, Hussain M. Effect of substituting fishmeal with black soldier fly and mealworm larvae meal on nutrient digestibility, body and mineral composition of Labeo rohita. Open Vet. J.. 2026; 16(1): 241-255. doi:10.5455/OVJ.2026.v16.i1.22 Web Style Ismat N, Hayat S, Hussain M. Effect of substituting fishmeal with black soldier fly and mealworm larvae meal on nutrient digestibility, body and mineral composition of Labeo rohita. https://www.openveterinaryjournal.com/?mno=280386 [Access: January 31, 2026]. doi:10.5455/OVJ.2026.v16.i1.22 AMA (American Medical Association) Style Ismat N, Hayat S, Hussain M. Effect of substituting fishmeal with black soldier fly and mealworm larvae meal on nutrient digestibility, body and mineral composition of Labeo rohita. Open Vet. J.. 2026; 16(1): 241-255. doi:10.5455/OVJ.2026.v16.i1.22 Vancouver/ICMJE Style Ismat N, Hayat S, Hussain M. Effect of substituting fishmeal with black soldier fly and mealworm larvae meal on nutrient digestibility, body and mineral composition of Labeo rohita. Open Vet. J.. (2026), [cited January 31, 2026]; 16(1): 241-255. doi:10.5455/OVJ.2026.v16.i1.22 Harvard Style Ismat, N., Hayat, . S. & Hussain, . M. (2026) Effect of substituting fishmeal with black soldier fly and mealworm larvae meal on nutrient digestibility, body and mineral composition of Labeo rohita. Open Vet. J., 16 (1), 241-255. doi:10.5455/OVJ.2026.v16.i1.22 Turabian Style Ismat, Nida, Sikandar Hayat, and Majid Hussain. 2026. Effect of substituting fishmeal with black soldier fly and mealworm larvae meal on nutrient digestibility, body and mineral composition of Labeo rohita. Open Veterinary Journal, 16 (1), 241-255. doi:10.5455/OVJ.2026.v16.i1.22 Chicago Style Ismat, Nida, Sikandar Hayat, and Majid Hussain. "Effect of substituting fishmeal with black soldier fly and mealworm larvae meal on nutrient digestibility, body and mineral composition of Labeo rohita." Open Veterinary Journal 16 (2026), 241-255. doi:10.5455/OVJ.2026.v16.i1.22 MLA (The Modern Language Association) Style Ismat, Nida, Sikandar Hayat, and Majid Hussain. "Effect of substituting fishmeal with black soldier fly and mealworm larvae meal on nutrient digestibility, body and mineral composition of Labeo rohita." Open Veterinary Journal 16.1 (2026), 241-255. Print. doi:10.5455/OVJ.2026.v16.i1.22 APA (American Psychological Association) Style Ismat, N., Hayat, . S. & Hussain, . M. (2026) Effect of substituting fishmeal with black soldier fly and mealworm larvae meal on nutrient digestibility, body and mineral composition of Labeo rohita. Open Veterinary Journal, 16 (1), 241-255. doi:10.5455/OVJ.2026.v16.i1.22 |

