




| Research Article | ||
Open Vet. J.. 2025; 15(12): 6738-6746 Open Veterinary Journal, (2025), Vol. 15(12): 6738-6746 Research Article Comparative morphometric analysis of Javan and Sunda slow lorises (Nycticebus spp.): Implications for species classification and body length estimationHery Wijayanto1*, Wendi Prameswari2, Nur Purba Priambada2, Ariana Ariana1, Woro Danur Wendo1 and Tri Wahyu Pangestiningsih11Department of Anatomy, Faculty of Veterinary Medicine, Universitas Gadjah Mada, Yogyakarta, Indonesia 2Yayasan Inisiasi Alam Rehabilitasi Indonesia, Bogor, Indonesia *Corresponding Author: Hery Wijayanto. Department of Anatomy, Faculty of Veterinary Medicine, Universitas Gadjah Mada, Yogyakarta, Indonesia. Email: herykh [at] ugm.ac.id Submitted: 02/07/2025 Revised: 15/11/2025 Accepted: 30/11/2025 Published: 30/11/2025 © 2025 Open Veterinary Journal
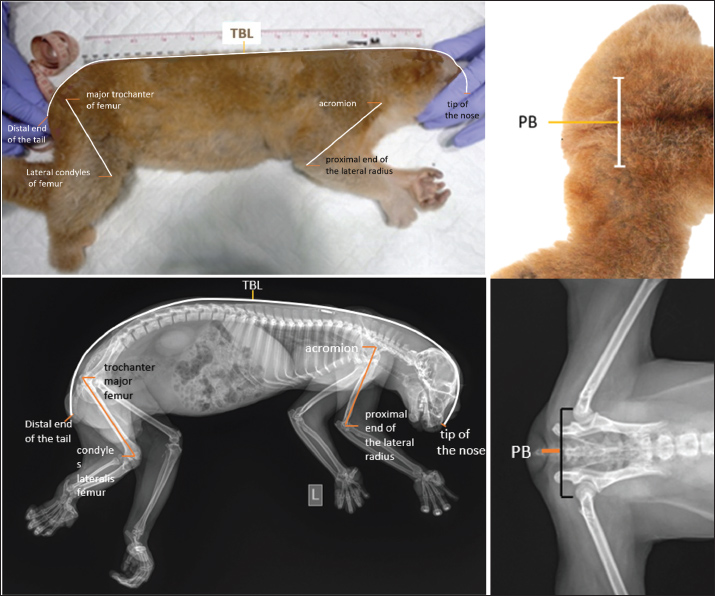
AbstractBackground: The islands of Sumatra and Java in Indonesia are home to slow lorises, and the long-standing biogeographical separation between the Sunda and Javan species contributes to morphological divergence. However, morphological identification and body size prediction remain limited due to insufficient anatomical data. Aim: This study aimed to investigate the anatomical characteristics of the thoracic and pelvic limbs in relation to the total body length (TBL) and pelvic breadth (PB) of the Sunda and Javan slow lorises, which are relevant to distinct biogeographic pressures. Methods: Morphometric data were collected from 43 adult slow lorises in a rehabilitation and conservation area under the supervision of Yayasan Inisiasi Alam Rehabilitasi Indonesia, Bogor. Measurements comprised the upper arm length, that considered the bony landmark of the humerus (HL), thigh length referring to the femur bone landmarks (FL), TBL, and PB. Measurements were taken during routine medical check-ups within a week. Statistical analyses, including correlation and regression, were conducted using the established methodologies. Results: The results showed that HL, FL, TBL, and PB of Javan slow lorises were significantly higher than those of Sunda, but there was no statistically significant difference between males and females. Both HL and FL were moderately correlated with TBL in both species, contributing to TBL prediction. HL and FL showed no significant correlation with PB. Conclusion: The body measurements of Sunda slow lorises were significantly smaller than those of the Javan species. These results provided useful morphometric data that improved species identification and helped estimate body length, but did not serve as reliable indicators of sexual dimorphism. The data obtained could help in comparative anatomy and forensic applications in conservation efforts, such as fur color pattern and body size identification. In line with genetic and ecological behavior data, the results are expected to serve as an important reference in conservation programs. Keywords: Javan slow lorises, Morphometric, Nycticebus coucang, Nycticebus javanicus, Sunda slow loris. IntroductionBiodiversity within a given habitat is intricately influenced by its region’s evolutionary and geological history. A region’s evolutionary and geological history intricately influences biodiversity within a given habitat. In Southeast Asia, the islands of Sumatra and Java, which have been geographically isolated for millions of years, present a compelling case of biogeographical divergence (Connell et al., 2019). It is hypothesized that this long-term separation has driven unique evolutionary trajectories and adaptive responses among the fauna of each island, contributing to the distinct biodiversity profiles (Leonard et al., 2015). Moreover, the separation caused adaptations that affected the morphology of animals in both islands, including slow lorises in Sunda and Javan (Arifin et al., 2022). These slow lorises are closely related species that diverged millions of years ago due to geological separation between the Sunda and Javan tectonic plates. Geographic and environmental factors, including temperature, prey availability, body size dynamics, and seasonal variation, continue to influence morphological differences between species (Nekaris et al., 2010; Blair et al., 2024). Comparative morphology and morphometric exploration of the Sunda and Javan Slow lorises are critically required to show the biogeographical characteristics of each species. In this context, morphological and morphometric studies have become essential tools in taxonomy, enabling the subtle detection and quantification of biologically meaningful differences between closely related taxa. Morphology refers to the form and structure of organisms, while morphometrics provides a quantitative framework for systematic exploration of these traits. These tools help identify characteristic features of species that can be obtained by genetic analysis. Slow lorises are nocturnal primates found in several Southeast Asian regions, including Indonesia. These species are distributed on the islands of Sumatra (Sunda slow lorises, Nycticebus coucang), Java (Javan slow lorises, Nycticebus javanicus), and Kalimantan (Bornean slow lorises, Nycticebus borneanus, Nycticebus menagensis). Due to habitat destruction, hunting, and illegal trade, the population is decreasing, causing the rescue of Sunda and Javan species in the rehabilitation center named Yayasan Inisiasi Alam Rehabilitasi Indonesia (YIARI), Bogor, Javan Island. The International Union for Conservation of Nature has listed the Sunda slow lorises (N. coucang) as an endangered species, while the Javan (N. javanicus) is a critically endangered species (Karimloo et al., 2023). In animal conservation, particularly in the rescue and rehabilitation from illegal trade, accurate species identification is essential to prevent misclassification and misplacement. Morphometrical and anatomical analyses of slow lorises (N. coucang and N. pygmaeus), including fossils of N. javanicus and N. coucang, show consistent patterns of clinal varying cranial and dental traits (Ravosa, 1998; Pozzi et al., 2015; Gruwier and Amano, 2023). Facial and skull morphology delineating species and subspecies in Bornean slow lorises N. menagensis, N. bancanus, N. borneanus, and Nycticebus kayan (Munds et al., 2013), and deep phylogenetic splits underpinning current diversity (Kongrit et al., 2020). These methods are essential for accurate species identification, understanding historical distributions, and uncovering cryptic lineages, key for conservation and taxonomy in slow lorises. Therefore, this study aimed to identify anatomical differences between Sunda and Javan slow lorises by analyzing limb proportions relative to TBL and PB. In conservation contexts, the results provided useful morphometric information for body size estimation, sexual dimorphism, species identification, comparative anatomy, and forensic applications. The data strongly emphasize evolutionary, biogeographical, or genetic aspects. Materials and MethodsPreparation of animals and body measurementsThis study was conducted with the support of the YIARI, Bogor, Indonesia, using 43 live adult slow lorises. The sample comprised 12 male and 19 female Javan slow lorises, as well as 5 male and 7 female Sunda slow lorises. However, limited information was obtained on the age of the animals, as Sunda and Javan slow lorises were rescued from illegal wildlife trade. Data were collected during routine medical check-ups under the supervision of a YIARI veterinarian within a week. General anesthesia was administered using zolazepam-tiletamine (Zoletil®, Virbac, France) at a dose of 5 mg/kg of body weight, delivered through intramuscular injection at the caudal extremities. Measurements were taken when the animals lay on a flat and horizontal surface in a supine position with the nose positioned perpendicular to the body axis. Total body length (TBL), upper arm length (HL), and thigh length (FL) were measured using a measuring tape, while hip or PB was determined with a calibrated digital caliper and measuring tape. All measurements were conducted following established osteometry guidelines (Welbourne and Clason, 1975; von den Driesch, 1976) by considering the following bony structures: 1. TBL is the sum of the head-body length and tail length. 2. The head and body length is the longest measurement from the tip of the nose to the base of the tail. Measured when the animal lies on a flat plane parallel to the back. 3. Tail length is the length of the tail from the proximal end of the cauda to the distal end of the tail and does not include hair. 4. PB is the distance between the right and left trochanter major. 5. The upper arm length (HL) is measured from the acromion to the proximal end of the lateral radius, with the arm positioned consistently. 6. Thigh length (FL) is measured from the most prominent point on the hip in the lateral part (trochanter major femur) to the lateral part of the knee (condyles lateralis femur), with the foot parallel to the tibial axis (Fig. 1).
Fig. 1. Body measurements of TBL, femur, humerus length, and PB. External body measurement (above) and X-ray image (below) (pictures courtesy of YIARI). Data analysisHL and FL were used as independent variables, whereas TBL and PB were analyzed separately as dependent variables. Three different observers independently collected morphometric data to minimize measurement errors. The mean values from the three sets of measurements were used for statistical analysis. Data were analyzed using IBM SPSS 26, which included normality tests (Kolmogorov–Smirnov), comparative (Mann–Whitney), correlation (Spearman’s), data transformation (z-score), and multiple linear regression. The Mann–Whitney test was used to analyze non-parametric samples of two independent variables, such as comparing males and females or interspecific Sunda and Javan of each morphometric parameter. The correlation coefficients were interpreted following previously described guidelines (Mukaka, 2012; Van Remoortel et al., 2020). Spearman’s correlation was considered appropriate for correlating continuous and non-parametric variables. Data normalization was performed using z-scores for regression analysis to standardize regression. Multiple linear regression was applied to predict TBL and PB based on HL and FL. The model for TBL was considered valid after confirming the normality of the residuals and the absence of significant multicollinearity between predictor variables. Generally, prediction models have explicitly accepted an R² value in the range of approximately 0.5–0.7 as being sufficiently accurate for predictive purposes. R² is a determination coefficient commonly used in regression analysis, showing the proportion of variance in the dependent variable that is explained by the independent variable(s). If r is the Pearson correlation coefficient, then R²=r². In simple linear regression, R² is just the square of the Pearson correlation coefficient (Pal and Lim, 1998; Roustaei, 2024). Ethical approvalThis study was approved by the Ethical Committee of the Faculty of Veterinary Medicine, Universitas Gadjah Mada (No. 002.0/EC-FKH/Int/2019). ResultsThe Sunda and Javan slow lorises exhibited morphological similarities in overall appearance. Specifically, Sunda slow lorises showed a broader white patch between the eyes, which widened across the frontal area, with limited white hair present on the lateral sides of the neck. Javan slow lorises are characterized by a distinct arrowhead-shaped patch of white hair between the eyes (interocular stripe), extending across the frontal region and often continuing along the neck to the dorsal thorax, interocular upper tip, and dorsal stripe extending to the back (Fig. 2).
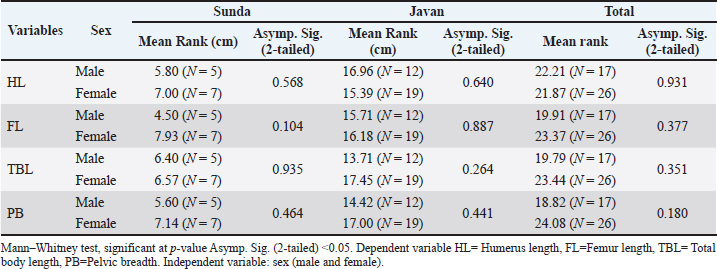
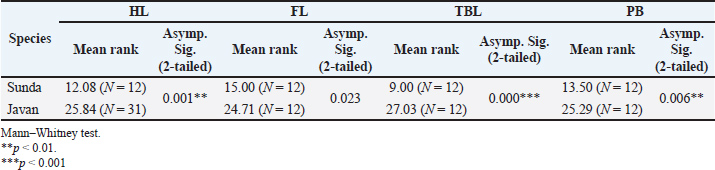
Fig. 2. Sunda slow lorises (A) show a broader white patch between the eyes (yellow arrow), which widens across the frontal area, with limited white hair on the lateral sides of the neck (red arrow). Javan slow lorises (B) are characterized by a distinct arrowhead-shaped patch of white hair between the eyes (interocular stripe) (yellow arrow), extending across the frontal region and often continuing along the neck to the dorsal thorax, interocular upper tip, and dorsal stripe extending to the back (red arrow) (YIARI). Measurement data (n=43) were assumed valid and reliable. The Kolmogorov–Smirnov test showed that the TBL was normally distributed (p > 0.05). However, HL, FL, and PB did not pass the normality test. Despite variable inhomogeneity, researchers used the Mann–Whitney nonparametric test for comparison. No statistically significant differences (p > 0.05) were observed when comparing HL, FL, TBL, and PB between males and females. This comparison was made considering the two grouped species. The parameters compared between males and females were not significant (p > 0.05), leading to further analysis within each Sunda and Javan species separately (Table 1). TBL was significantly different (p=0.000) between the two species without considering the sex. Significant differences in HL, FL, and PB were found between the Sunda and Javan species (p < 0.05; Table 2). The mean rank showed a larger size of HL, FL, TBL, and PB of Javan slow lorises compared with those of Sunda. Table 1. The males and females of each dependent variable in each species were compared separately, and the two species were grouped together to confirm sexual dimorphism.
Table 2. Comparison between the Sunda and Javan species without considering sex in each morphometric parameter of the study.
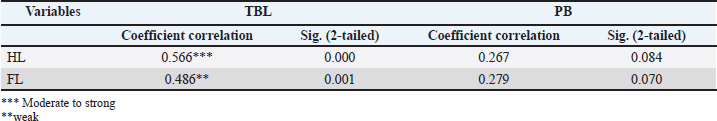
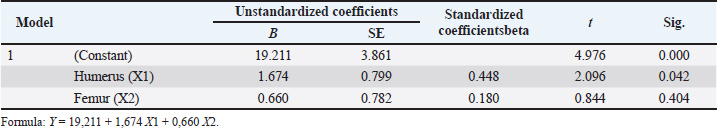
The Spearman correlation test showed a positive correlation between HL and TBL as well as between FL and TBL in all samples, without considering species and sex (p < 0.05). The correlation of HL to TBL was moderate to strong according to previous studies (Mukaka, 2012; Van Remoortel et al., 2020), whereas the correlation of FL to TBL was weak (rho) with values of 0.56 and 0.49, respectively (Table 3). There was no significant relationship (p > 0.05) between HL and FL and PB, while the correlation coefficient was low (Table 3). Therefore, HL could be used to predict TBL in both Sunda and Javan slow lorises. HL and FL were not considered useful in the PB prediction model due to low correlation. In this context, TBL prediction could be obtained using a regression model, which started with a normality test (Kolmogorov–Smirnov) for aggregate distribution. When the data were normal, the parametric linear regression test was used for analysis. HL and FL play a 36% role in estimating TBL (Table 4), with a 64% gap for other factors showing a significant contribution. The prediction model was obtained by the regression test as Y=19.211 + 1.674 X1 + 0.660 X2 for TBL (Table 5), considering X1 and X2 as HL and FL, respectively. Table 3. Spearman’s correlation test of upper arm length (HL), thigh length (FL), TBL, and PB.
Table 4. Prediction model (Dependent variable: TBL).
Table 5. Regression model (Dependent variable: TBL).
DiscussionMorphological evolution in every species often interacts with both biotic and abiotic environments (Jablonski, 2008). Although evolution also occurs at the genetic level, initial adaptation due to environmental pressures (such as climate change) will be preceded by morphological changes, such as body size in the population, due to animal plasticity (Merilä and Hendry, 2014). The physical differences between the Javan and the Sunda slow lorises are still very minimal despite the long-term separation. Therefore, the accurate identification of Sunda and Javan slow lorises is essential and must be conducted with caution to prevent the misplacement of rescued individuals, as these two species are closely related. Based on the average body measurements conducted in this study, the Javan slow lorises were larger than the Sunda. These results agreed with previous observations, where N. javanicus tended to show relatively greater overall body size than N. coucang (Nekaris and Jaffe, 2007). However, the differences were not found to be statistically significant, as the sample sizes were limited. Nekaris and Jaffe (2007) used 34 live slow lorises of 3 species (N. coucang, N. menagensis, and N. javanicus), which were rescued from the wildlife trade in Java, Indonesia. In this study, 43 samples were used, comprising 12 male and 19 female Javan, as well as 5 male and 7 female Sunda slow lorises. The results showed significant differences in HL, FL, TBL, and PB, with the Javan species being larger than the Sunda species. The geographical isolation of Java and Sumatra Island for millions of years could be responsible for the differences in size (Leonard et al., 2015; Hinckley et al., 2020). Species must adapt to survive in a particular environment, and over long evolutionary timeframes, adaptations can lead to morphological changes (Graham et al., 2017). The longer limb of Javan slow lorises compared to Sunda is predicted to occur due to differences in habitat quality of the lower vegetation on Java. This condition makes slow Javan lorises roam more widely for the same nutritional value. According to Wolff’s Law, the bone in a healthy person or animal will adapt and remodel in response to the loads and forces placed on it. This provides that a more active limb will increase in length and girth (Thompson, 2012). Morphological evolution represents the adaptive response of an animal to its environment because of natural selection and genetic drift. Adaptation includes the prey habits and food sources in each region, which contribute to morphological divergence among species (Burrows et al., 2015; Natasya et al., 2025). Sumatra Island possesses less natural biomass, including food for slow lorises that are primarily exudativorous, leading to feeding on insects, flower nectar, sap, fruit, and small vertebrates (Burrows et al., 2015). In this study, the physical variations represent long-term evolutionary reactions to the unique ecological circumstances of each island. Island syndrome refers to a set of phenotypic traits observed in species, such as reduced body size or altered behavior, due to unique ecological pressures (Millien, 2006). These traits often arise from factors such as reduced predation, lower biodiversity, and relaxed competition. For instance, species on islands can evolve to smaller sizes due to the absence of large predators and limited resources, leading to adaptations that differ from those of mainland relatives. A craniometric study of slow lorises (Nycticebus spp.) in Southeast Asia reported that the cranium size of Javan (N. javanicus) was larger than that of the Sunda species (N. coucang). The smallest cranial dimensions were observed in the Bornean slow lorises (N. menagensis) (Groves and Maryanto, 2008). Similarly, this study showed that the body dimensions of the Javan slow lorises were larger than those of the Sunda. In rescue centers, the risk of accidental hybridization increases because of the housing of multiple species and individuals from illegal trade (Nekaris and Jaffe, 2007). Hybridization could impact or alter the genetic integrity of populations of slow lorises. Additionally, human intervention in the care and treatment can cause complications of health problems (Nekaris and Jaffe, 2007), which influence growth and body size. The correlation of HL with TBL is moderate but not significant with PB. Similar to the anthropometric correlation model, limb length is predominantly correlated with stature, whereas PB is related to body weight (Hartono, 2018). A recent study reported that hip circumference was the most significantly correlated variable with changes in body weight and condition score in slow lorises (Ghassani et al., 2023). Observations have also shown that a larger hip circumference could be caused by fat deposit thickness (Meguro et al., 2015; Huo et al., 2022). Slow lorises generally show specialized PB adaptations that correlate with slow and suspensory locomotion. These traits are functionally convergent with xenarthrans and show evolutionary trade-offs for arboreal efficiency (Lewton and Dingwall, 2016). Studies of the morphology and morphometrics of the pelvic membrane ossa in small mammals, such as the Javan tree-shrew (Tupaia javanica), plain squirrels (Callosciurus notatus), and sugar gliders (Petaurus breviceps), have shown sexual dimorphism in the coxae, where the pubis is wider in females than in males (unpublished data). A wider ossa coxae in females contributes to increased PB, which influences species measurements depending on the sample’s sex distribution. However, this study did not provide results of sexual dimorphism in SL in the Sunda and Javan based on PB, as statistical analysis showed no significant differences between males and females. Previous reports on sexual dimorphism between slow lorises showed little to no pronounced sexual dimorphism in size or weight between males and females in species such as Sunda. Both sexes are generally similar in terms of coloration and body size (Doherty, 2024). This study provides TBL prediction based on limb proportion. The model can be acceptable because long limb bones, including the humerus and femur, are common proxies as skeletal predictors, but there is distinct allometry between humans and non-human primates (Hens et al., 1998). Tarsal bones provide more accurate predictions of body length and mass in Eocene primates than dental measurements (Dagosto and Terranova, 1992). However, this study has limitations in body weight measurements and age, considering only length evaluations. The results indicate that adult animals with different body condition scores do not show differences in bone length. The evaluation is not based on the weight parameter but on the length measurements. The heavier animals do not show larger bone size because the body conditions of the sample used vary based on the feeding pattern. Furthermore, body weight can still vary with fat deposits (Meguro et al., 2015; Huo et al., 2022), causing inaccurate results. Previous studies have often shown that length increases rapidly with age, followed by a gradual decrease until some asymptote or plateau occurs when the animal reaches maturity (de Santiago et al., 2015; Xu et al., 2023). This study only used adult mature animals for morphometric analysis. The TBL prediction model is obtained from moderate to strong Spearman’s correlation by R² values (57%) of HL. In a previous study, all dependent variables had good predictive accuracy with moderate R² values (Wijaya et al., 2022). A systematic review of the prediction model for medical usage rates in mass gatherings evaluated the model fit using R², which ranged from 0% (no fit) to 100% (perfect fit). Predictive performance was categorized as very weak (0%–4%), weak (4%–16%), moderate (16%–36%), strong (36%–64%), or very strong (64%–100%) (Van Remoortel et al., 2020). Therefore, the model of TBL prediction in this study was accepted from HL and FL, which showed 49% of R² values because both limb proportions considered have a strong correlation. This study establishes a solid scientific foundation with clear biological and conservation relevance. The results address the economic and commercial importance of biodiversity management, particularly in regions of wildlife conservation, ecotourism, and biodiversity management in Indonesia and Southeast Asia. The data obtained strengthen species-specific identification, which is essential for accurate record-keeping, tracking, and management in both rehabilitation centers and wildlife reserves by providing reliable morphometric data for distinguishing between Sunda and Javan slow lorises. The ability to differentiate species with greater precision allows for more targeted conservation planning. This is because species misidentification can lead to the improper allocation of limited conservation resources, thereby reducing the effectiveness of rehabilitation programs. Reliable morphometric tools ensure that rescued species are correctly classified, managed, and released into appropriate habitats, thereby increasing survival rates and reducing post-release monitoring or recapture costs. Moreover, accurate species classification is essential in combating illegal wildlife trade. Promoting scientifically verified distinctions between endemic species, such as Sunda and Javan slow lorises, can enhance biodiversity narratives and conservation messaging in the ecotourism and education sectors. Enhanced clarity and differences among species can increase public interest and foster more support from ecotourism stakeholders and conservation funding organizations. ConclusionIn conclusion, this study shows that PB, HL, FL, and TBL differ significantly between Sunda and Javan slow lorises, but no substantial variation is observed between males and females. The slow Sunda lorises also have significantly smaller body dimensions. HL and FL are moderately correlated with TBL in both species, contributing to the prediction of OL, although PB shows no significant relationship. These results provide valuable morphometric data that enhance species identification and body length estimation. However, these data are not reliable indicators for assessing sexual dimorphism. This is because differences in morphology between Javan and Sunda slow lorises play an important role in comparative anatomy and forensic conservation, as rescue centers often rely on physical traits for initial identification. Applications in the forensic field can be used for species identification by determining the relationship between confiscated bone parts based on the correlation between HL, FL, and TBL. AcknowledgmentsThe authors are grateful to the Dean of the Faculty of Veterinary Medicine, Universitas Gadjah Mada, Yayasan Inisiasi Alam Rehabilitasi Indonesia (YIARI), Bogor, Indonesia, and Heribertus for the amazing photographs of Javan and Sunda slow lorises. The authors also thank Richard Stephen Moore for a detailed review of the manuscript. Conflict of interestThe authors declare no conflict of interest. FundingThe authors declare that this study received no specific grant. Author’s contributionsHW supervised the study design and manuscript. WP, WDW, AR, PNP, and TWP were responsible for sample collection, data analysis, and manuscript drafting. All authors have read and approved the final version of the manuscript. Data availabilityThe original data and contributions from this study are available in the article or supplementary materials. Further inquiries should be directed to the corresponding author. ReferencesArifin, U., Smart, U., Husemann, M., Hertwig, S.T., Smith, E.N., Iskandar, D.T. and Haas, A. 2022. Phylogeographic inference of Sumatran ranids bearing gastromyzophorous tadpoles concerning the Pleistocene drainage systems of Sundaland. Scientific. Rep. 12(1), 1–12; doi:10.1038/s41598-022-14722-9 Blair, M.E., Cao, G.T.H., López-Nandam, E.H., Veronese-Paniagua, D.A., Birchette, M.G., Kenyon, M., Md-Zain, B.M., Munds, R.A., Nekaris, K.A.I., Nijman, V., Roos, C., Thach, H.M., Sterling, E.J. and Le, M.D. 2024. Correction to: molecular Phylogenetic Relationships and Unveiling Novel Genetic Diversity among Slow and Pygmy Lorises, including Resurrection of Xanthonycticebus intermedius (Genes, 2023). Genes 15(4), 15; doi:10.3390/genes15040451 Burrows, A.M., Hartstone-Rose, A. and Nash, L.T. 2015. Exudativory in the Asian loris, Nycticebus: evolutionary divergence in the toothcomb and M3 species. Am. J. Phys. Anthropology 158(4), 663–672; doi:10.1002/ajpa.22829 Connell, K.A.O., Smart, U., Sidik, I., Riyanto, A., Kurniawan, N., Smith, E. N., Worth, F., & Connell, K.A.O. 2019. Diversification of bent-toed geckos ( Cyrtodactylus ) on Sumatra and West Java. Department of Vertebrate Zoology and Global Genome Initiative , Museum of Natural History , Smithsonian Institute , Washington , District of Columbia , 20560 , USA. Dagosto, M. and Terranova, C.J. 1992. Estimating the body size of eocene primates: a comparison of dental and postcranial variables. Int. J. Primatology 13(3), 307–344; doi:10.1007/BF02547818 De Santiago, H.A.R., De Pierro, L.R., Reis, R.M., Caluz, A.G., Ribeiro, V.B. and Volpon, J.B. 2015. Allometric relationships among body mass, MUZZLE-tail length, and tibia length during growth in Wistar rats. Acta. Cirurgica. Brasileira. 30(11), 743–748; doi:10.1590/S0102-865020150110000004 Doherty, L. 2024. Characterization of slow loris species using shape-based three-dimensional cranial geometric morphometrics Ghassani, Y., Rianti, P., Priambada, N.P., Arifin, I., Saptorini, I., Prameswari, W. and Darusman, H. 2023. Welfare assessment of slow loris (Nycticebus spp.) at an Indonesian primate rehabilitation center: body condition score development and validation. Am. J. Primatology 85, e23524; doi:10.1002/ajp.23524 Graham, N.R., Gruner, D.S., Lim, J.Y. and Gillespie, R.G. 2017. Island Ecology and Evolution: challenges in the Anthropocene. Environ. Conservation. 44(4), 323–335; doi:10.1017/S0376892917000315 Groves, C. and Maryanto, I. 2008. Craniometry of slow lorises (genus Nycticebus) in insular Southeast Asia Primates of the Oriental Night, 115–122. Gruwier, B. and Amano, N. 2023. Holocene record of the slow loris (Nycticebus sp.) in Java, Indonesia. Am. J. Biol. Anthropology. 182(3), 467–475; doi:10.1002/ajpa.24840 Hartono. and M. 2018. Indonesian anthropometry update for special populations revisited: drillis and Contini. Int. J. Ind. Ergonom. 64, 89–101; doi:10.1016/j.ergon.2018.01.004 Hens, S.M., Konigsberg, L.W. and Jungers, W.L. 1998. Estimation of the African ape body length. J. Hum. Evol. 34(3), 401–411. Hinckley, A., Hawkins, M.T.R., Achmadi, A.S., Maldonado, J.E. and Leonard, J.A. 2020. Ancient Divergence Driven by Geographic Isolation and Ecological Adaptation in Sundaland Tree Squirrels. Front. Ecol. Evol. 8, 1–18; doi:10.3389/fevo.2020.00208 Huo, D.L., Bao, M.H., Cao, J. and Zhao, Z.J. 2022. Cold exposure prevents fat accumulation in striped hamsters refed a high-fat diet following food restriction. BMC. Zool. 7(1), 1–12; doi:10.1186/s40850-022-00122-z Karimloo, L., Campera, M., Imron, M.A., Rakholia, S., Mehta, A., Hedger, K. and Nekaris, K.A.I. 2023. Habitat Use, Terrestriality and Feeding Behavior of Javan Slow Lorises in Urban Areas of a Multi-Use Landscape in Indonesia. Land 12(7), 1–18; doi:10.3390/land12071349 Kongrit, C., Markviriya P Laithong. and Khudamrongsawat. 2020. Species Identification and Unlocking of Confiscated Slow Lorises (Nycticebus spp.) Based on the Mitochondrial DNA Markers. Folia Primatologica 91(1), 1–14; doi:10.1159/000500007 Leonard, J.A., Den Tex, R.J., Hawkins, M.T.R., Muñoz-Fuentes, V., Thorington, R. and Maldonado, J.E. 2015. Phylogeography of vertebrates on the Sunda Shelf: a multi-species comparison. J. Biogeography. 42(5), 871–879; doi:10.1111/jbi.12465 Lewton, K.L. and Dingwall, H.L. 2016. Morphological convergence in the pubis of slow-moving primates and xenarthrans: a systematic review. Am. J. Phys. Anthropology 161(3), 381–397; doi:10.1002/ajpa.23038 Meguro, S., Hasumura, T. and Hase, T. 2015. Body fat accumulation in zebrafish is induced by a diet rich in fat and reduced by supplementation with green tea extract. PLos One. 10(3), 1–12; doi:10.1371/journal.pone.0120142 Merilä, J. and Hendry, A.P. 2014. Climate change, adaptation, and phenotypic plasticity: the problem and evidence. Evol. Applications. 7(1), 1–14; doi:10.1111/eva.12137 Millien. 2006. Morphological evolution is accelerated in island mammals. PLos Biol. 4(10), 1863–1868; doi:10.1371/journal.pbio.0040321 Mukaka, M.M. 2012. Statistics corner: a guide to the appropriate use of correlation coefficients in medical research. Malawi. Med. J. 24(3), 69–71. Munds, R.A., Nekaris. and Ford. 2013. Taxonomy of the Bornean Slow Loris with the New Species Nycticebus kayan (Primates, Lorisidae). Am. J. Primatology 75(1), 46–56; doi:10.1002/ajp.22071 Natasya, A.M., Rianti, P., Darusman, H.S. and Widiyani, T. Feeding behavior and food palatability of Sunda slow loris (Nycticebus coucang) at the IPB University Primates Research Center. In Bioinformatics Web of Conferences, Les Ulis, France, EDP Sciences, 2025, 162, pp 1–10; doi: 10.1051/bioconf/202516200002 Nekaris, K.A.I. and Jaffe, S. 2007. Unexpected diversity of slow lorises (Nycticebus spp.) within the Javan pet trade: implications for slow loris taxonomy. Contributions. To. Zool. 76(3), 187–196; doi:10.1163/18759866-07603004 Nekaris, K.A.I., Shepherd, C.R., Starr, C.R. and Nijman, V. 2010. Exploring cultural drivers for wildlife trade via an ethnoprimatological approach: a case study of slender and slow lorises (Loris and Nycticebus) in South and Southeast Asia. Am. J. Primatology. 72(10), 877–886. Pal, N. and Lim, W.K. 1998. On the multiple determination coefficient in a linear regression model. J. Italian Stat. Soc. 7(2), 129–157; doi:10.1007/bf03178925 Pozzi, L., Nekaris A Perkin., Bearder ER Pimley., Schulze U Streicher., Nadler A Kitchener., Zischler D Zinner. and Roos. 2015. Ancient divergences among neglected lorisiform primates. Zoological J. Linnean Soc. 175(3), 661–674; doi:10.1111/zoj.12286 Ravosa, M.J. 1998. Cranial allometry and geographic variation in slow lorises. Am. J. Primatology 45(3), 225–243. Roustaei, N. 2024. Application and interpretation of linear regression analysis. Med. Hypothesis. Discovery. Innov. Ophthalmology 13(3), 151–159; doi:10.51329/mehdiophthal1506 Van Remoortel, H., Scheers, H., De Buck, E., Haenen, W. and Vandekerckhove, P. 2020. Prediction modeling studies for medical usage rates in mass gatherings: a systematic review. PLos One 15(6), 1–20; doi:10.1371/journal.pone.0234977 von den Driesch, A. 1976. A guide to the measurement of animals bones from archaeological sites, Peabody Museum of Archaeology and Ethnology, Cambridge, MA: Harvard University. Welbourne, R.G. and Clason, A.T. 1975. Archaeozoological Studies. In The South African Archeological Bulletin (Vol. 30, Issue 119/120). https://doi.org/10.2307/3888103 Wijaya, T.T., Jiang, P., Mailizar, M. and Habibi, A. 2022. Predicting Factors Influencing Preservice Teachers’ Behavior Intention in STEM Education Implementation Using Partial Least Squares Approach. Sustainability. (Switzerland). 14(16), 14; doi:10.3390/su14169925 Xu, W., Feng, Y., Gu, Z., Zhang, S., Yang, Z., Xu, Y. and Gu, H. 2023. Effects of morphological traits on body weight and analysis of growth-related genes of Parabramis pekinensis at different ages. BMC. Zool. 8(1), 1–11; doi:10.1186/s40850-023-00174-9 | ||


| How to Cite this Article |
| Pubmed Style Wijayanto H, Prameswari W, Priambada NP, Ariana A, Wendo WD, Pangestiningsih TW. Comparative morphometric analysis of Javan and Sunda slow lorises (Nycticebus spp.): Implications for species classification and body length estimation. Open Vet. J.. 2025; 15(12): 6738-6746. doi:10.5455/OVJ.2025.v15.i12.53 Web Style Wijayanto H, Prameswari W, Priambada NP, Ariana A, Wendo WD, Pangestiningsih TW. Comparative morphometric analysis of Javan and Sunda slow lorises (Nycticebus spp.): Implications for species classification and body length estimation. https://www.openveterinaryjournal.com/?mno=268260 [Access: January 25, 2026]. doi:10.5455/OVJ.2025.v15.i12.53 AMA (American Medical Association) Style Wijayanto H, Prameswari W, Priambada NP, Ariana A, Wendo WD, Pangestiningsih TW. Comparative morphometric analysis of Javan and Sunda slow lorises (Nycticebus spp.): Implications for species classification and body length estimation. Open Vet. J.. 2025; 15(12): 6738-6746. doi:10.5455/OVJ.2025.v15.i12.53 Vancouver/ICMJE Style Wijayanto H, Prameswari W, Priambada NP, Ariana A, Wendo WD, Pangestiningsih TW. Comparative morphometric analysis of Javan and Sunda slow lorises (Nycticebus spp.): Implications for species classification and body length estimation. Open Vet. J.. (2025), [cited January 25, 2026]; 15(12): 6738-6746. doi:10.5455/OVJ.2025.v15.i12.53 Harvard Style Wijayanto, H., Prameswari, . W., Priambada, . N. P., Ariana, . A., Wendo, . W. D. & Pangestiningsih, . T. W. (2025) Comparative morphometric analysis of Javan and Sunda slow lorises (Nycticebus spp.): Implications for species classification and body length estimation. Open Vet. J., 15 (12), 6738-6746. doi:10.5455/OVJ.2025.v15.i12.53 Turabian Style Wijayanto, Hery, Wendi Prameswari, Nur Purba Priambada, Ariana Ariana, Woro Danur Wendo, and Tri Wahyu Pangestiningsih. 2025. Comparative morphometric analysis of Javan and Sunda slow lorises (Nycticebus spp.): Implications for species classification and body length estimation. Open Veterinary Journal, 15 (12), 6738-6746. doi:10.5455/OVJ.2025.v15.i12.53 Chicago Style Wijayanto, Hery, Wendi Prameswari, Nur Purba Priambada, Ariana Ariana, Woro Danur Wendo, and Tri Wahyu Pangestiningsih. "Comparative morphometric analysis of Javan and Sunda slow lorises (Nycticebus spp.): Implications for species classification and body length estimation." Open Veterinary Journal 15 (2025), 6738-6746. doi:10.5455/OVJ.2025.v15.i12.53 MLA (The Modern Language Association) Style Wijayanto, Hery, Wendi Prameswari, Nur Purba Priambada, Ariana Ariana, Woro Danur Wendo, and Tri Wahyu Pangestiningsih. "Comparative morphometric analysis of Javan and Sunda slow lorises (Nycticebus spp.): Implications for species classification and body length estimation." Open Veterinary Journal 15.12 (2025), 6738-6746. Print. doi:10.5455/OVJ.2025.v15.i12.53 APA (American Psychological Association) Style Wijayanto, H., Prameswari, . W., Priambada, . N. P., Ariana, . A., Wendo, . W. D. & Pangestiningsih, . T. W. (2025) Comparative morphometric analysis of Javan and Sunda slow lorises (Nycticebus spp.): Implications for species classification and body length estimation. Open Veterinary Journal, 15 (12), 6738-6746. doi:10.5455/OVJ.2025.v15.i12.53 |

