




| Research Article | ||
Open Vet. J.. 2026; 16(5): 2619-2630 Open Veterinary Journal, (2026), Vol. 16(5): 2619-2630 Research Article Nano-formulated curcumin restores hypothalamic–pituitary–ovarian axis disturbance in cadmium-induced female reproductive toxicityMohammed Nabeel Abbas Al-Muhja1 and Jabbar Abbas Ahmed Al-Saaidi2*1Department of Physiology, Pharmacology, and Biochemistry, College of Veterinary Medicine, University of Al-Qadisiyah, Al Diwaniyah, Iraq 2Department of Basic Sciences, College of Dentistry, Al-Ayen Iraqi University, Nasiriyah, Iraq *Corresponding Author: Jabbar Abbas Ahmed Al-Saaidi. Department of Basic Sciences, College of Dentistry, Al-Ayen Iraqi University, Nasiriyah, Iraq. Email: jabbar.alsaaidi [at] alayen.edu.iq Submitted: 18/02/2026 Revised: 14/04/2026 Accepted: 23/04/2026 Published: 31/05/2026 © 2025 Open Veterinary Journal
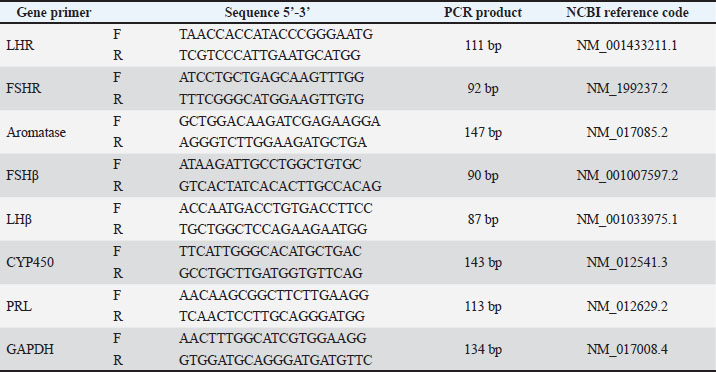
AbstractBackground: Cadmium (Cd) is a pervasive environmental toxicant associated with female reproductive dysfunction through endocrine disruption and oxidative stress. Strategies that enhance antioxidant bioavailability may improve therapeutic efficacy against heavy metal-induced ovarian damage. Aim: To investigate the protective efficacy of polycaprolactone/polyvinyl alcohol-loaded curcumin nanoparticles (PCL/PVA/Cm NPs) against CdCl₂-induced reproductive toxicity in female rats, focusing on hormonal modulation and hypothalamic–pituitary–ovarian axis gene expression. Methods: Forty-eight adult female rats were allocated into six groups: control, CdCl₂, Cm, CdCl₂ + Cm, PCL/PVA/Cm NPs, and CdCl₂+PCL/PVA/Cm NPs. Body and reproductive organ weight, Serum gonadotropin-releasing hormone (GnRH), follicle stimulating-hormone (FSH), luteinizing hormone (LH), estradiol, progesterone, and prolactin (PRL) levels were assessed after 4 weeks. Ovarian (LHR, FSHR, and aromatase) and pituitary (LHβ, FSHβ, CYP450, and PRL) gene expression levels were quantified. Results: The results revealed that CdCl₂ exposure significantly reduced body, ovarian, and uterine weights and suppressed GnRH, FSH, LH, estradiol, and progesterone levels while elevating PRL levels. Ovarian and pituitary reproductive gene expression was markedly downregulated. Cm partially reversed these effects; however, nanoformulated Cm produced significantly greater restoration, normalizing hormonal profiles and gene expression patterns to near-control values. Conclusion: The nano-encapsulation markedly enhances Cm bioefficacy, effectively restoring endocrine balance and molecular signaling within the reproductive axis. PCL/PVA/Cm NPs represent a promising nanotherapeutic strategy for mitigating Cd-induced female reproductive toxicity. Keywords: Cadmium chloride, Curcumin nanoparticles, Endocrine disruption, Polycaprolactone, Polyvinyl alcohol. IntroductionCadmium (Cd) is a common environmental and industrial contaminant that has a long biological half-life and easily accumulates in soft tissues, such as reproductive organs. Cd is a strong endocrine disruptor and reproductive toxicant in females because it affects body growth, ovarian and uterine structure, and hormonal homeostasis (Genchi et al., 2020; Hu et al., 2023). Experimental and epidemiological evidence has shown that Cd exposure modifies body weight gain and organ weights by disrupting metabolism and altering adipose tissue function, mostly through oxidative stress- and inflammatory-mediated pathways (Attia et al., 2022; Liu et al., 2023). In female rodents, subchronic exposure to Cd decreases ovarian and uterine weight, disrupts estrous cyclicity, and causes histopathological lesions, such as follicular atresia and stromal degeneration (Nasiadek et al., 2019; Quddus et al., 2021). The ovary is susceptible to Cd-induced oxidative damage owing to its high vascularity and steroidogenic function. Cd produces reactive oxygen species (ROS), damages mitochondrial function, and initiates apoptotic signaling pathways, resulting in granulosa cell death and folliculogenesis failure (Zhang et al., 2020; Hu et al., 2023). Cd disrupts the hypothalamic–pituitary–gonadal (HPG) axis, by inhibiting of gonadotropin synthesis and release from the pituitary. Changes in the gene expression of the pituitary luteinizing hormone (LH) and follicle stimulating-hormone (FSH) were reported in response to Cd exposure, mainly as a consequence of oxidative stress and estrogen-like activities (Saedi et al., 2022; Santiago-Andres et al., 2025). Furthermore, Cd negatively affects ovarian steroidogenesis by altering the activity of critical enzymes and signaling pathways, such as P450-dependent systems, which can lead to alterations in serum estradiol and progesterone levels (Zhao et al., 2021; Sheweita et al., 2023). Molecularly, Cd exposure reduces the expression of genes involved in follicle development and survival, resulting in exhausted ovarian reserve and compromised follicular maturation (Goutam Mukherjee et al., 2022; Xiao and Lai, 2025). Considering that oxidative stress plays a key role in Cd-induced reproductive toxicity, the antioxidant-mediated therapies are of great interest. Curcumin (Cm), the major polyphenolic compound in Curcuma longa, has attracted attention as a candidate natural phytochemical owing to its powerful antioxidant, anti-inflammatory, and anti-apoptotic effects (Aggarwal and Harikumar, 2009; Hewlings and Kalman, 2017). Cm also scavenges ROS and recruits endogenous antioxidant enzymes and redox-sensitive signaling pathways, including Nrf2/HO-1 and PI3K/Akt (Menon and Sudheer, 2007; Wang et al., 2024). Cm exerted Protective effects on Cd-induced oxidative damage in ovarian tissue and preserved the normal architecture around follicles and hormonal balance (Yang et al., 2019; Flory et al., 2021). Furthermore, Cm is known to modulate reproductive endocrine function by affecting the levels of gonadotropin secretion and steroidogenic gene expression by supporting folliculogenesis and ovarian reserve maintenance (Ghosh et al., 2015; Lv et al., 2021). Cm’s clinical efficacy is hindered by its poor aqueous solubility, rapid metabolism, and low systemic bioavailability (Wahlström and Blennow, 1978; Anand et al., 2007). To address these factors, Cm nanoformulations such as polymeric nanoparticles (NPs), lipid-based nanocarriers, and nanoemulsions, have been synthesized, which have been reported to greatly improve their stability, absorption, and tissue distribution (Yallapu et al., 2012; Yadav et al., 2012). Cm NPs have stronger antioxidant and anti-inflammatory effects than native Cm and show better protection from metal induction toxicity (Sarawi et al., 2022; Rashwan et al., 2022). Recent studies have shown that nano Cm can enhance the ovarian follicle count, stabilize serum sexual hormones, and modulate gene expression implicated in the activation and survival pathways of follicles (Virk et al., 2025; Hendarto et al., 2025). Additionally, nanoparticlate-based delivery systems improve the potential of Cm in modulating the pituitary–gonadal axis signaling and counteracting oxidative stress-induced apoptosis in genital tissues (Yadav et al., 2012; Nazarian et al., 2025). Although there have been independent investigations related to the reproductive toxicity induced by Cd or protective roles of Cm and its nanocomposites, integrated studies that jointly consider changes in body weight, ovarian/uterine weights, serum reproductive hormone profile, pituitary and ovarian gene expression modulations, as well as a detailed evaluation of ovarian folliculogenesis, are available only at a limited level. Interestingly, no extensive molecular-histological correlations have been performed to date concerning nano-Cm treatment of the HPG axis gene and follicular dynamics modulation on Cd exposure. Thus, the current experiment was designed with an aim to bridge this important gap by systematically reassessing the ameliorative potential of Cm nanocomposites against Cd-induced female reproductive toxicity at physiological, hormonal, molecular, and histomorphometric levels. It may also lay scientific groundwork for the formulation of nanophytotherapeutic methods to alleviate heavy metal-mediated reproductive impairments. Materials and MethodsExperimental animalsForty-eight adult female Wistar rats weighing 180–220 g were procured from the animal house of the institute and kept in standard laboratory conditions (temperature, 22℃ ± 2°C; light/dark cycle, 12 hours each; relative humidity, 50%–60%) for 1 week before the initiation of the experiment. The rats were provided with free access to a standard pellet diet and water. Rats were randomly divided into six experimental groups (n=8 per group): T1, negative control; T2, CdCl₂ 30 mg/kg/day) (El-Demerdash et al., 2004); T3, Cm 50 mg/kg/day) (Tarlan et al., 2025); T4, CdCl₂ + Cm; T5, polycaprolactone (PCL)/polyvinyl alcohol (PVA)/Cm NPs 0.207 mg/kg/day; and T6, CdCl₂ + NPs. The treatments were orally administered for 4 weeks. The experimental design followed OECD guideline 407 for a repeated-dose oral toxicity study (Toyoda et al., 2000). Body weight and relative organ weight measurementA digital balance was used to measure the initial and final body weights. Body weight gain was measured as the difference between the final and initial body weight. Ovarian and uterine weights were expressed relative to body weight as follows: (organ weight/final body weight) × 100. Body and organ weights have been employed as indices of systemic and reproductive toxicity (Toyoda et al., 2000; Faroon et al., 2012). Hormonal assaysCardiac puncture was used to obtain blood samples, which were then centrifuged for serum retrieval. Serum gonadotropin-releasing hormone (GnRH), FSH, LH, prolactin (PRL), estradiol, and progesterone were measured in serum using commercial enzyme-linked immunosorbant assay kits (SUNLONG COMPANY, CHINA) according to the manufacturer’s instructions. Such hormone determinations were made to evaluate the integrity of the hypothalamic–pituitary–ovarian axis, which has been shown to be perturbed by Cd exposure (Zhang et al., 2020; Goutam Mukherjee et al., 2022). Molecular studies of ovarian and pituitary gene expressionThe ovarian and pituitary tissues were dissolved in TRIzol reagent, and total RNA was prepared as described by Chomczynski and Sacchi (1987). RNA concentration and purity were determined spectrophotometrically. Complementary DNA was generated from 1 µg RNA. Transcription levels of ovarian LHR, FSHR, and aromatase (CYP19A1) genes and pituitary FSHβ, LHβ, CYP450, and PRL genes were measured by quantitative real-time PCR (qRT-PCR) with SYBR Green chemistry. The 2−ΔΔCt method was used to calculate relative gene expression with β-actin as a housekeeping gene (Livak and Schmittgen, 2001). These are the vital control genes of folliculogenesis and steroidogenesis (Richards and Pangas, 2010). Primer designThe primers used in this study were the GAPDH gene primer as a housekeeping gene, 1) ovarian LHR, FSHR, and aromatase genes; and 2) pituitary FSHβ, LHβ, CYP450, and PRL genes as target genes for gene expression analysis (Table 1). These primers were designed using the National Center for Biotechnology Information Gene Bank database and Primer 3 design online. The primers were used in the quantification of gene expression levels using the qRT-PCR technique based on SYBR Green DNA-binding dye, which was supported by Bioneer Co. (Korea). Table 1. Primers used in this study.
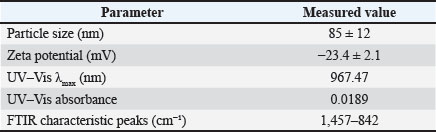
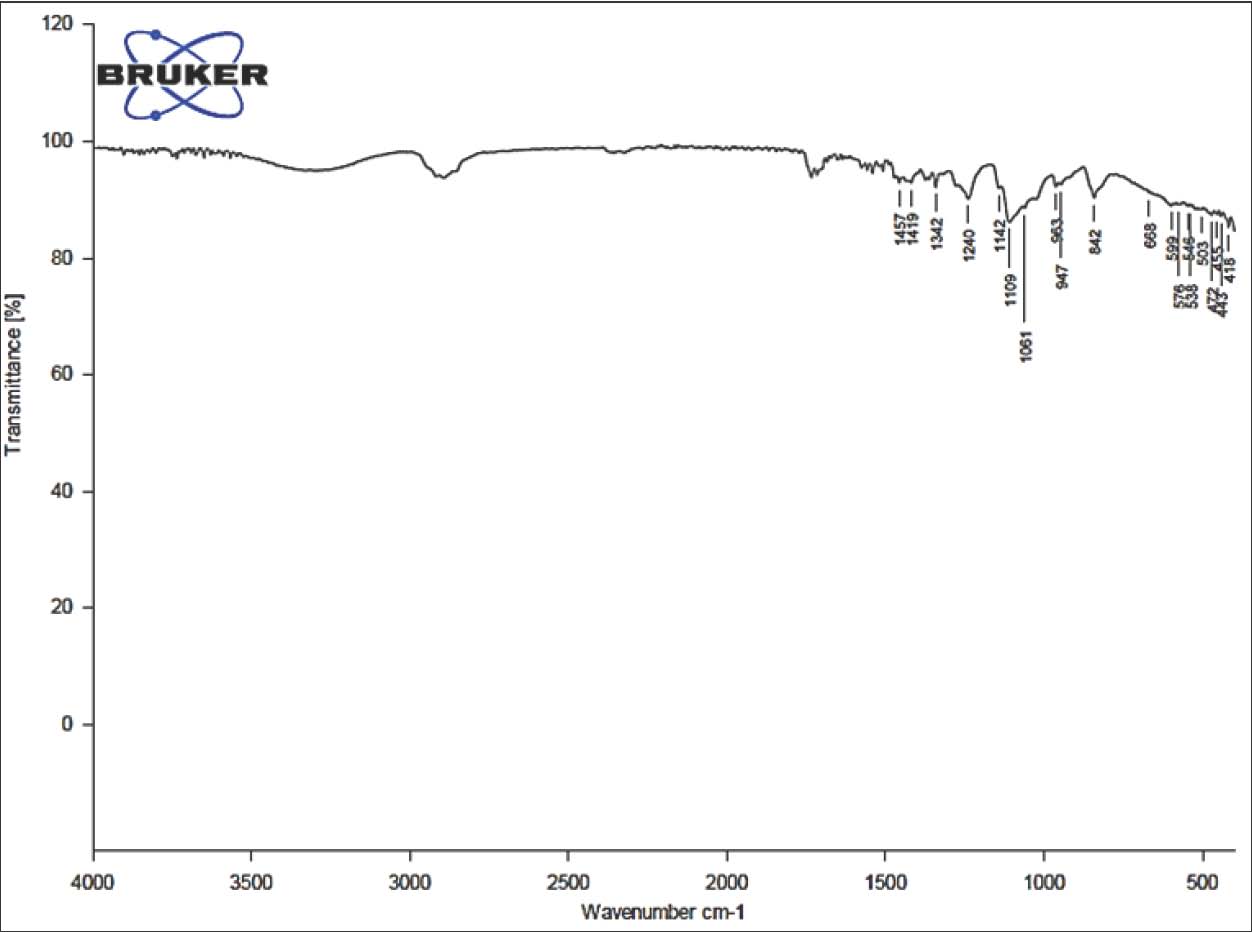
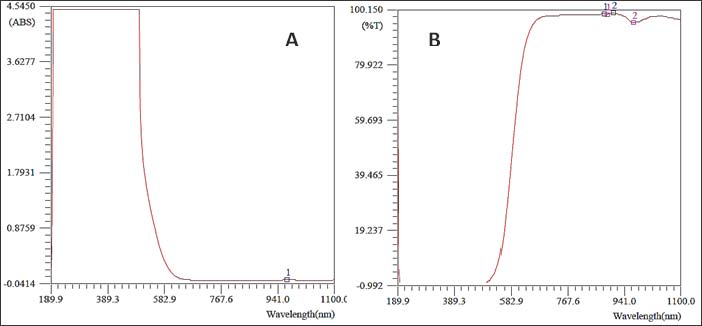
Statistical analysisValues are presented as mean ± SEM. Data were analyzed statistically by one-way analysis of variance followed by Tukey’s post hoc test. All statistical analysis were performed, and p-value <0.05 was considered statistically significant (Snedecor and Cochran, 1989). Ethical approvalAll animal experiments were conducted in accordance with guidelines established by the National Institutes of Health for the care and use of laboratory animals (National Research Council (US) Committee for the Update of the Guide for the Care and Use of Laboratory Animals, 2011). This study was approved by the ethics and policy committee of the College of Veterinary Medicine, Al-Qadisiyah University (No. 1180). ResultsPhysical characterization of the PCL/PVA/Cm NPsTable 2 summarizes the physical characteristics of the nanocomposite PCL/PVA/Cm NPs with a particle size of 85 ± 12. UV–Vis spectrophotometric analysis demonstrated a characteristic absorption peak (λmax) at 967.47 nm with an absorbance value of 0.0189, confirming the successful incorporation of Cm within the polymeric matrix (Fig. 1). The high transmittance observed across the visible and near-infrared regions indicated good NP suspension dispension stability and optical clarity (Fig. 2). FTIR spectroscopy further verified NP formation and drug encapsulation. Characteristic absorption bands corresponding to PCL, PVA, and Cm were detected at 1,457, 1,419, 1,342, 1,240, 1,142, 1,109, 1,061, 963, 947, and 842 cm⁻¹ (Fig. 3A). Detailed inspection of the fingerprint region (Fig. 3B) confirmed the preservation of functional groups without evidence of chemical degradation, indicating stable polymer-drug interactions. Table 2. Physical characterization parameters of PCL, PVA, Cm NPs.
Fig. 1. UV–Visible absorption spectrum of PCL/PVA/Cm NPs showing the characteristic optical response and λmax.
Fig. 2. UV–Visible transmittance (%T) spectrum of the PCL/PVA/Cm NPs.
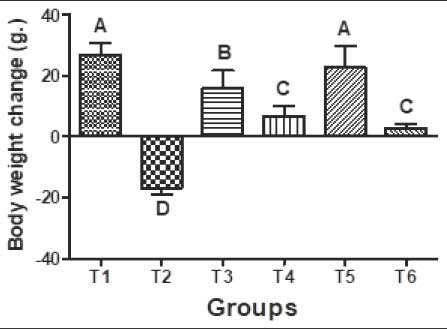
Fig. 3. A) FTIR spectrum of PCL/PVA /Cm NPs illustrating characteristic functional groups. B) Enlarged FTIR fingerprint region of PCL/PVA/Cm NPs highlighting key absorption bands. Body weight and relative ovarian and uterine weights changesAs illustrated in Figure 4, significant differences (p < 0.05) in body weight were observed among all experimental groups after 4 weeks of treatment. The control group (T1) showed a consistent increase in body weight, whereas the Cd-treated group (T2) exhibited a marked reduction in body weight compared with the other groups (p < 0.05). Cm-treated animals (T3) demonstrated significant improvement compared with the Cd group, although values remained slightly lower than those of the control. Co-treatment with CdCl₂ and Cm (T4) significantly improved body weight compared with T2. Notably, the PCL/PVA/Cm NP group (T5) exhibited greater weight gain than the free Cm group. Similarly, CdCl₂ + PCL/PVA/Cm NPs (T6) significantly restored body weight compared with T2, with values approaching those of T4.
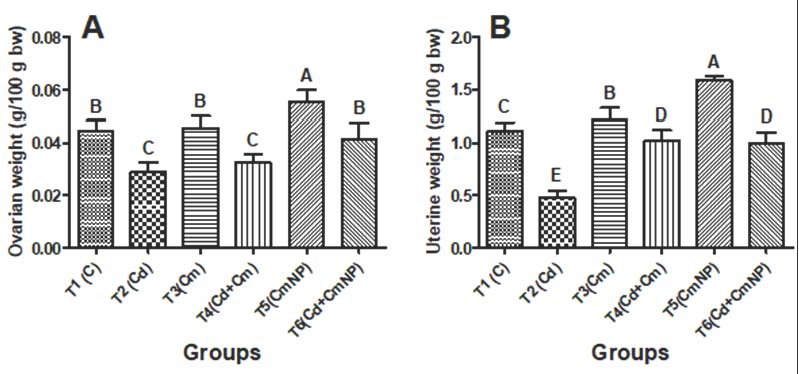
Fig. 4. Body weight changes (g.) of female rats after 4 weeks of supplementation with distilled water (T1), CdCl2 (T2), Cm (T3), combination of CdCl2 and Cm (T4), PCL/PVA/Cm NPs (T5), and combination of CdCl2 and PCL/PVA/Cm combination (T6). Values are presented as mean ± standard deviation. Different letters denote significant differences (p < 0.05) between groups. Changes in ovarian and uterine weights are presented in Figure 5A and B, respectively. Exposure to Cd (T2) significantly reduced ovarian weight compared with control (T1). Cm alone (T3) maintained an ovarian weight comparable to that of the control. However, CdCl₂ + Cm (T4) failed to fully restore the ovarian mass. Treatment with PCL/PVA/Cm NPs alone (T5) resulted in increased ovarian weight, while CdCl₂ + PCL/PVA/Cm NPs (T6) restored ovarian mass close to control values. Similarly, the uterine weight was significantly decreased in the Cd-treated group. Cm alone significantly elevated uterine weight compared with the control group. The CdCl₂ + Cm group showed partial recovery, whereas the T5 group exhibited the highest uterine weight among all groups. Combined CdCl₂ + PCL/PVA/Cm NPs (T6) improved uterine weight relative to Cd alone.
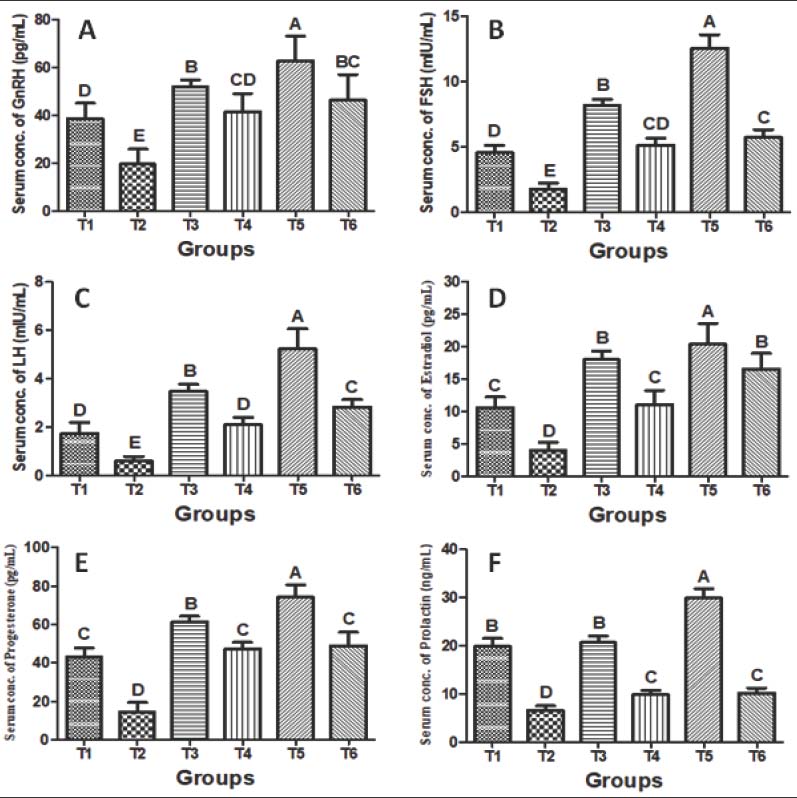
Fig. 5. Relative ovarian (A) and uterine (B) weights (g/100 g of body weight) of female rats after 4 weeks of supplementation with distilled water (T1), CdCl2 (T2), Cm (T3), a combination of CdCl2 and Cm (T4), PCL/PVA/Cm NPs (T5), and a combination of CdCl2 and PCL/PVA/Cm NPs (T6). Values are presented as mean ± standard deviation. Different letters denote significant differences (p < 0.05) between groups. Serum hormonal profileSerum hormonal changes are depicted in Fig. 6A–F. Exposure to Cd significantly reduced serum GnRH (Fig. 6A), FSH (Fig. 6B), LH (Fig. 6C), estradiol (Fig. 6D), progesterone (Fig. 6E), and PRL (Fig. 6F) levels compared with the control (p < 0.05). Cm treatment (T3) significantly enhanced most hormonal parameters compared with those in the Cd group. Co-treatment (T4) partially restored hormonal concentrations toward normal values. Cm-loaded NPs (T5) produced the most pronounced hormonal enhancement across all parameters. Moreover, CdCl₂ + PCL/PVA/Cm NPs (T6) markedly restored hormone levels compared with Cd alone and demonstrated superior protective efficacy compared with free Cm cotreatment.
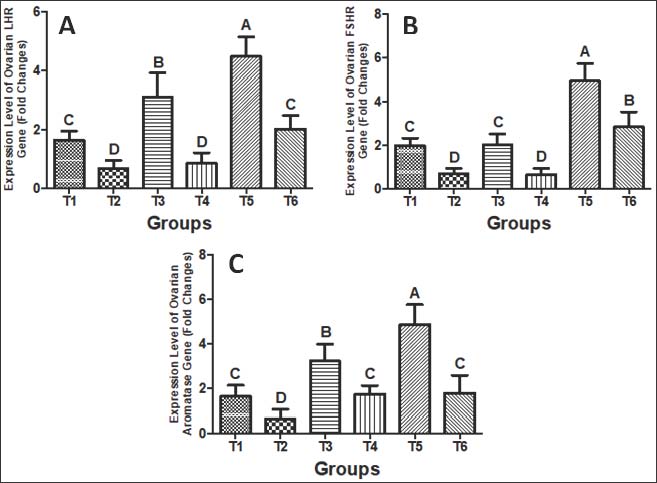
Fig. 6. Serum levels of: A) GnRH (pg/ml), B) FSH (mIU/ml), C) LH (mIU/ml), D) estradiol (pg/ml), E) progesterone (pg/ml), and F) PRL (ng/ml) in female rats after 4 weeks of treatment with distilled water (T1), CdCl2 (T2), Cm (T3), combination of CdCl2 and Cm (T4), PCL/PVA/Cm NPs (T5), and combination of CdCl2 and PCL/PVA/Cm NPs (T6). Values are presented as mean ± standard deviation. Different letters denote significant differences (p < 0.05) between groups. Ovarian gene expressionThe relative mRNA expression levels of the ovarian LH receptor, FSH receptor, and aromatase genes are shown in Figure 7A–C. All three genes were significantly downregulated by Cd chloride compared to the control (p < 0.05). Cm (T3) and PCL/PVA/Cm NPs (T5) significantly upregulated gene expression relative to T2. While CdCl₂ + Cm (T4) demonstrated limited recovery, CdCl₂ + PCL/PVA/Cm NPs (T6) exhibited marked restoration of gene expression, indicating enhanced protective efficacy of the nanoformulation.
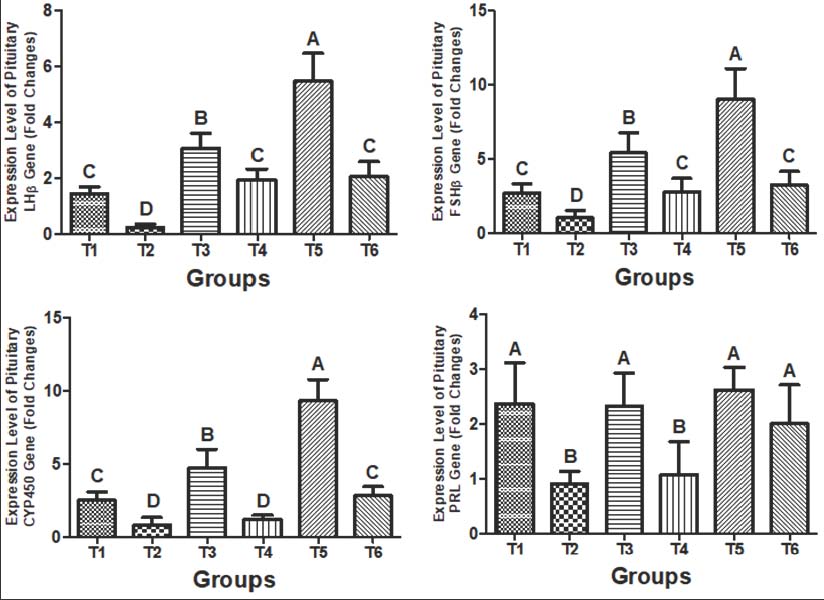
Fig. 7. Gene expression levels (fold changes) of: A) LH receptor, B) FSH receptor, and C) aromatase in the ovarian tissue of female rats after 4 weeks of treatment with distilled water (T1), CdCl2 (T2), Cm (T3), combination of CdCl2 and Cm (T4), PCL/PVA/Cm NPs (T5), and combination of CdCl2 and PCL/PVA/Cm combination (T6). Values are presented as mean ± standard deviation. Different letters denote significant differences (p < 0.05) between groups. Pituitary gene expressionThe expression patterns of pituitary LHβ, FSHβ, CYP450, and PRL genes are illustrated in Figure 8A–D. Exposure to Cd significantly suppressed all measured genes (p < 0.05). Cm and NP treatments significantly elevated gene expression compared with the Cd group. The T5 group showed the highest upregulation, particularly for CYP450 (Figs. 4–8C). Co-treatment with CdCl₂ and PCL/PVA/Cm NPs (T6) demonstrated stronger restoration than co-treatment with free Cm (T4).
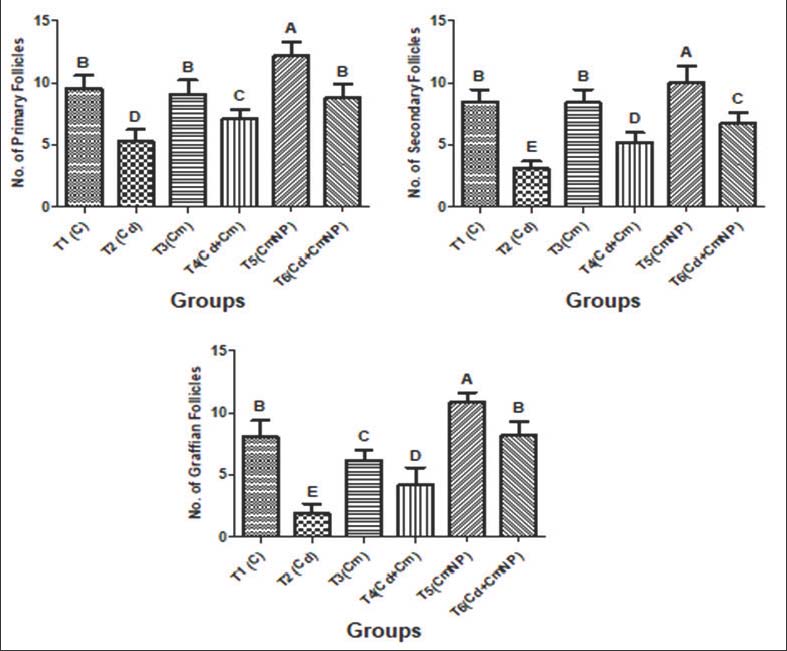
Fig. 8. Gene expression levels (fold changes) of: A) LHβ, B) FSHβ, C) CYP450, and D) PRL in the pituitary tissue of female rats after 4 weeks of treatment with distilled water (T1), CdCl2 (T2), Cm (T3), combination of CdCl2 and Cm (T4), PCL/PVA/Cm NPs (T5), and combination of CdCl2 and PCL/PVA/Cm NPs (T6). Values are presented as mean ± standard deviation. Different letters denote significant differences (p < 0.05) between groups. Follicular countFigure 9 shows the quantitative assessment of primary, secondary, and Graafian follicles. After 4 weeks of treatment, the control group exhibited a normal and balanced number. There was a significant decrease in the number of primary, secondary, and Graafian follicles following exposure to CdCl₂ compared to controls (p < 0.05). The administration of Cm only sustained primary and secondary follicles at the same level as that in the control group, whereas it caused a significant reduction in the number of Graafian follicles (p < 0.05). The combined application of CdCl₂ and Cm also led to a marked reduction in all types of follicles (p < 0.05). However, follicle numbers at each developmental stage were significantly higher in Cm-loaded PCL/PVA NPs treated groups (p < 0.05). CdCl₂, along with Cm-loaded NPs, CdCl2 resulted in primary and Graafian follicles similar to controls, whereas secondary follicles were still repressed.
Fig. 9. Number of ovarian follicles (primary, secondary, and Graffian follicles) of female rats after 4 weeks of supplementation with distilled water (T1), CdCl2 (T2), Cm (T3), combination of CdCl2 and Cm (T4), PCL/PVA/Cm NPs (T5), and combination of CdCl2 and PCL/PVA/Cm NPs (T6). Values are presented as mean ± standard deviation. Different letters denote significant differences (p < 0.05) between groups. DiscussionIn the current study, we established that CdCl2 exposure evoked significant reproductive toxicity in female rats as indicated by the loss of weight gain, decrease in ovarian and uterine relative weights, and imbalance of serum hormonal profile (FSH/LH/estradiol), which further led to downregulation of expression genes in both the ovary and pituitary, along with declined follicular numbers. Significantly, Cm (especially in its PCL/PVA nanocomposite form) exhibited a remarkable ability to reduce these reproductive impairments, suggesting the improved therapeutic potency of nano Cm for HPG axoprotection. Cd is a classical endocrine-disrupting chemical that displays toxicity through oxidative stress, inflammation, steroidogenesis, and gonadotropin signaling inhibition (Genchi et al., 2020; Hu et al., 2023). Subchronic exposure to Cd decreases body weight and perturbs metabolic homeostasis (Kunnumakkara et al., 2017; Liu et al., 2023). The recovery of body weight after Cm and nano-Cm treatment can be attributed to strong antioxidant and anti-inflammatory effects (Hewlings and Kalman, 2017; Wang et al., 2024). In addition, nano Cm formulations further increase the systemic bioavailability and tissue distribution compared with free Cm (Yadav et al., 2012; Das and Al-Naemi, 2019). The significant reduction in ovarian and uterine weights in Cd-treated rats aligns with previous findings demonstrating Cd-induced ovarian atrophy and uterine degeneration (Priyadarsini, 2014; Nasiadek et al., 2019; Xiao and Lai, 2025). Cm supplementation mitigates ovarian oxidative stress and preserves ovarian morphology (Yang et al., 2019; Park et al., 2021). Importantly, nano-Cm exhibited greater restoration of ovarian and uterine mass, likely due to improved cellular uptake and sustained release properties (Das and Al-Naemi, 2019; Rashwan et al., 2022). Changes in serum hormonal concentrations, such as the decrease in GnRH, FSH, LH, estradiol, and progesterone or increase in PRL levels, indicate that Cd can affect the HPG axis at various levels (Goutam Mukherjee et al., 2022; Santiago-Andres et al., 2025). Cm effectively reversed these hormonal parameters by intervening in the oxidative stress mechanisms and controlling steroidogenic enzymes (Moniruzzaman and Min, 2020; Wang et al., 2024). Nano Cm showed better hormone recovery than free Cm (Karthikeyan et al., 2020; Virk et al., 2025). At the molecular level, Cd downregulates the ovarian LH receptor, FSH receptor, and aromatase genes as well as pituitary LHβ, FSHβ, CYP450, and PRL genes (Saedi et al., 2022; Goutam Mukherjee et al., 2022). Cm also possesses an antioxidative effect and induces the catytochrome P450 system together with other antioxidant defense systems (Zhao et al., 2021; Silvestre et al., 2023) that can restore steroidogenic equilibrium. The nanoformulation exhibited more potent gene upregulation, indicating enhanced intracellular delivery. Follicular counts showed that Cd had an adverse effect by decreasing the number in primary, secondary, and Graafian follicles. Heavy metals that act through oxidative stress and apoptosis compromise folliculogenesis (Hu et al., 2023; Xiao and Lai, 2025). Cm protects the ovarian reserve by triggering the P13K/Akt and Nrf2/HO-1 pathways (Duan et al., 2024; Hendarto et al., 2025). Nano-Cm also significantly improves the number of follicles compared with native Cm (Virk et al., 2025; Hendarto et al., 2025). Altogether, nano-Cm was more effective in preventing Cd-induced reproductive toxicity than ordinary Cm. Through the regulation of the balance, gene expression, follicular development-related hormones, and genes, PCL/PVA/Cm NPs had a good effect on Cd-induced destruction of the HPG axis. ConclusionIn summary, Cd chloride exposure induced marked reproductive toxicity in female rats by disturbing the HPG axis, which manifested as decreased body and relative reproductive organ weights, hormonal imbalance, decreased expression of key pituitary and ovarian genes, and disturbance of folliculogenesis. These adverse effects could be counterbalanced by Cm treatment, which acts as a regulator of steroidogenesis and an antioxidant. Subsequently, the hormone level, gene expression, and ovarian structure were compensated. Remarkably, the PCL/PVA nano Cm formulation offered better protective activity than free Cm, presumably due to significant increases in bioavailability and cellular uptake. These observations indicate that nano Cm is a potential combat substance for protecting the female reproductive system from Cd toxicity. AcknowledgmentThe authors acknowledge the Deanery of the College of Veterinary Medicine, University of Al-Qadisiyah, Iraq. Conflict of interestThe authors declare no conflict of interest. FundingThis research received no specific grant. Authors’ contributionBoth authors have contributed to writing, design, compilation, and editing of the final manuscript. Data availabilityAll data supporting this study’s finding are available within the manuscript and will be provided to the editor upon request. ReferencesAggarwal, B.B. and Harikumar, K.B. 2009. Potential therapeutic effects of curcumin, the anti-inflammatory agent, against neurodegenerative, cardiovascular, pulmonary, metabolic, autoimmune and neoplastic diseases. Interna. J. Biochem. Cell. Biol. 41(1), 40–59; doi:10.1016/j.biocel.2008.06.010 Anand, P., Kunnumakkara, A.B., Newman, R.A. and Aggarwal, B.B. 2007. Bioavailability of curcumin: problems and promises. Mol. Pharm. 4(6), 807–818; doi:10.1021/mp700113r Attia, S.M., Das, S.C., Varadharajan, K. and Al-Naemi, H.A. 2022. White adipose tissue as a target for cadmium toxicity. Front. Pharmacol. 13, 1010817; doi:10.3389/fphar.2022.1010817 Chomczynski, P. and Sacchi, N. 1987. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 162(1), 156–159; doi:10.1006/abio.1987.9999 Das, S.C. and Al-Naemi, H.A. 2019. Cadmium toxicity: oxidative stress, inflammation and tissue injury. Occup. Dis. Environ. Med. 7, 144–163; doi:10.4236/odem.2019.74012 Duan, H., Yang, S., Yang, S., Zeng, J., Yan, Z., Zhang, L., Ma, X., Dong, W., Zhang, Y., Zhao, X., Hu, J. and Xiao, L. 2024. The mechanism of curcumin to protect mouse ovaries from oxidative damage by regulating AMPK/mTOR mediated autophagy. Phytomed. Int. JPphytoth. Phytopharmacol. 128, 155468; doi:10.1016/j.phymed.2024.155468 El-Demerdash, F.M., Yousef, M.I., Kedwany, F.S. and Baghdadi, H.H. 2004. Cadmium-induced changes in lipid peroxidation, blood hematology, biochemical parameters and semen quality of male rats: protective role of vitamin e and beta-carotene. Food. Chem. Toxicol. 42, 1563–1571; doi:10.1016/j.fct.2004.05.001 Faroon, O., Ashizawa, A., Wright, S., Tucker, P., Jenkins, K., Ingerman, L. and Rudisill, C. 2012. Toxicological profile for cadmium. Atlanta, GA: Agency for Toxic Substances and Disease Registry (US). Flory, S., Männle, R. and Frank, J. 2021. The inhibitory activity of curcumin on P-glycoprotein and its uptake by and efflux from LS180 cells is not affected by its galenic formulation. Antioxidants 10(11), 1826; doi:10.3390/antiox10111826 Genchi, G., Sinicropi, M.S., Lauria, G., Carocci, A. and Catalano, A. 2020. The effects of cadmium toxicity. Int. J. Environ. Res. Public Health 17(11), 3782; doi:10.3390/ijerph17113782 Ghosh, S., Banerjee, S. and Sil, P.C. 2015. The beneficial role of curcumin on inflammation, diabetes and neurodegenerative disease: a recent update. Food. Chem. Toxicol. 83, 111–124; doi:10.1016/j.fct.2015.05.022 Goutam Mukherjee, A., Ramesh Wanjari, U., Renu, K., Vellingiri, B. and Valsala Gopalakrishnan, A. 2022. Heavy metal and metalloid - induced reproductive toxicity. Environ. Toxicol. Pharmacol. 92, 103859; doi:10.1016/j.etap.2022.103859 Hendarto, H., Hutama, S.A., Primariawan, R.Y., Alkaff, F.F., Utomo, B.S., Widjiati, W. and Suzuki, N. 2025. Nano-curcumin potentially ameliorates hormonal function and follicular counts following the vitrification and transplantation of rat ovarian tissue. Sci. Rep. 15(1), 34588; doi:10.1038/s41598-025-18081-z Hewlings, S. and Kalman, D. 2017. Curcumin: a review of its effects on human health. Foods 6(10), 92; doi:10.3390/foods6100092 Hu, B., Liu, S., Luo, Y., Pu, J., Deng, X., Zhou, W., Dong, Y., Ma, Y., Wang, G., Yang, F., Zhu, T. and Zhan, J. 2023. Procyanidin B2 alleviates uterine toxicity induced by cadmium exposure in rats: the effect of oxidative stress, inflammation, and gut microbiota. Ecotoxicol. Environ. Saf. 263, 115290; doi:10.1016/j.ecoenv.2023.115290 Karthikeyan, A., Senthil, N. and Min, T. 2020. Nanocurcumin: a promising candidate for therapeutic applications. Front. Pharmacol. 11, 487; doi:10.3389/fphar.2020.00487 Kunnumakkara, A.B., Bordoloi, D., Padmavathi, G., Monisha, J., Roy, N.K., Prasad, S. and Aggarwal, B.B. 2017. Curcumin, the golden nutraceutical: multitargeting for multiple chronic diseases. Brit. J. Pharmacol. 174(11), 1325–1348; doi:10.1111/bph.13621 Liu, X., Qi, M., Li, X., Wang, J. and Wang, M. 2023. Curcumin: a natural organic component that plays a multi-faceted role in ovarian cancer. J. Ovarian Res. 16(1), 47; doi:10.1186/s13048-023-01120-6 Livak, K.J. and Schmittgen, T.D. 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25(4), 402–408; doi:10.1006/meth.2001.1262 Lv, Y., Cao, R.C., Liu, H.B., Su, X.W., Lu, G., Ma, J.L. and Chan, W.Y. 2021. Single-oocyte gene expression suggests that curcumin can protect the ovarian reserve by regulating the PTEN-AKT-FOXO3a pathway. Int. J. Mol. Sci. 22(12), 6570; doi:10.3390/ijms22126570 Menon, V.P. and Sudheer, A.R. 2007. Antioxidant and anti-inflammatory properties of curcumin. Adv. Exp. Med. Biol. 595, 105–125; doi:10.1007/978-0-387-46401-5_3 Moniruzzaman, M. and Min, T. 2020. Curcumin, curcumin nanoparticles and curcumin nanospheres: a review on their pharmacodynamics based on monogastric farm animal, poultry and fish nutrition. Pharmaceutics 12(5), 447; doi:10.3390/pharmaceutics12050447 Nasiadek, M., Danilewicz, M., Klimczak, M., Stragierowicz, J. and Kilanowicz, A. 2019. Subchronic exposure to cadmium causes persistent changes in the reproductive system in female wistar rats. Oxid. Med. Cell. Longev. 2019, 6490820; doi:10.1155/2019/6490820 National Research Council (US) Committee for the Update of the Guide for the Care and Use of Laboratory Animals. 2011. Guide for the care and use of laboratory animals, Nazarian, M., Aramjoo, H., Roshanravan, B., Samarghandian, S. and Farkhondeh, T. 2025. Protective effects of curcumin and nanomicelle curcumin on chlorpyrifos-induced oxidative damage and inflammation in the uterus, ovary and brain of rats. Curr. Pharm. Biotech. 26(4), 490–496; doi:10.2174/0113892010297408240319073735 Park, J.H., Lee, B.M. and Kim, H.S. 2021. Potential protective roles of curcumin against cadmium-induced toxicity and oxidative stress. J. Toxicol. Environ. Health. Part. B. Crit. Rev. 24(3), 95–118; doi:10.1080/10937404.2020.1860842 Priyadarsini, K. 2014. The chemistry of curcumin: from extraction to therapeutic agent. Molecules 19(12), 20091–20112; doi:10.3390/molecules191220091 Quddus, A., Yimer, N., Jesse, F.F.A., Basit, M.A., Amir, M. and Islam, M.S. 2021. Edible bird's nest protects histomorphology of rat's uterus against cadmium (Cd) toxicity through a reduction of Cd deposition and enhanced antioxidant activity. Saudi J. Biol. Sci. 28(12), 7068–7076; doi:10.1016/j.sjbs.2021.08.012 Rashwan, A.K., Karim, N., Xu, Y., Hanafy, N.A.N., Li, B., Mehanni, A.H.E., Taha, E.M. and Chen, W. 2022. An updated and comprehensive review on the potential health effects of curcumin-encapsulated micro/nanoparticles. Crit. Rev. Food Sci. Nutr. 63(29), 9731–9751; doi:10.1080/10408398.2022.2070906 Richards, J.S. and Pangas, S.A. 2010. The ovary: basic biology and clinical implications. J. Clin. Invest. 120(4), 963–972; doi:10.1172/JCI41350 Saedi, S., Namavar, M.R., Shirazi, M.R.J., Rezazadeh, F.M. and Tsutsui, K. 2022. Exposure to Cadmium alters the population of glial cell types and disrupts the regulatory mechanisms of the HPG Axis in prepubertal female rats. Neurotoxicol. Res. 40(4), 1029–1042; doi:10.1007/s12640-022-00516-4 Santiago-Andres, Y., Hernández Álvarez, E., Ochoa Gutierrez, D., Morton Bermea, O. and Fiordelisio, T. 2025. In vivo effects of cadmium on signaling and secretion of pituitary gonadotrophs in male mice are time-dependent. J. Endocrinol. 267(1), e250161; doi:10.1530/JOE-25-0161 Sarawi, W.S., Alhusaini, A.M., Fadda, L.M., Alomar, H.A., Albaker, A.B., Alghibiwi, H.K., Aljrboa, A.S., Alotaibi, A.M., Hasan, I.H. and Mahmoud, A.M. 2022. Nano-curcumin prevents copper reproductive toxicity by attenuating oxidative stress and inflammation and improving Nrf2/HO-1 signaling and pituitary-gonadal axis in male rats. Toxics 10(7), 356; doi:10.3390/toxics10070356 Sheweita, S.A., Rafea, A.A.E. and Elbana, S.G. 2023. The deleterious effects of cadmium on oxidative stress markers, drug-metabolizing, and antioxidant enzyme activities: role of Silymarin and garlic as antioxidants. Environ. Sci. Pollut. Res. Int. 30(52), 112490–112502; doi:10.1007/s11356-023-30197-1 Silvestre, F., Santos, C., Silva, V., Ombredane, A., Pinheiro, W., Andrade, L., Garcia, M., Pacheco, T., Joanitti, G., Luz, G. and Carneiro, M. 2023. Pharmacokinetics of curcumin delivered by nanoparticles and the relationship with antitumor efficacy: a systematic review. Pharmaceuticals 16(7), 943; doi:10.3390/ph16070943 Snedecor, G.W. and Cochran, W.G. 1989. Statistical methods, 8th ed. Ames, IA: State University Press. Tarlan, M., Moradi, S., Heidarizade, N., Tavallaei, O., Khazayel, S., Farzaei, M.H. and Echeverría, J. 2025. The protective role of curcumin in mitigating drug-induced toxicity in male reproductive system. Front. Pharmacol. 16, 1620732; doi:10.3389/fphar.2025.1620732 Toyoda, K., Shibutani, M., Tamura, T., Koujitani, T., Uneyama, C. and Hirose, M. 2000. Repeated dose (28 days) oral toxicity study of flutamide in rats, based on the draft protocol for the 'Enhanced OECD Test Guideline 407' for screening for endocrine-disrupting chemicals. Arch. Toxicol. 74(3), 127–132; doi:10.1007/s002040050664 Virk, T.L., Liu, Q., Yuan, Y., Xu, X. and Chen, F. 2025. Curcumin as therapeutic modulator of impaired antioxidant defense system: implications for oxidative stress-associated reproductive dysfunction. Biology 14(7), 750; doi:10.3390/biology14070750 Wahlström, B. and Blennow, G. 1978. A study on the fate of curcumin in the rat. Acta Pharmacol. Toxicol. 43(2), 86–92; doi:10.1111/j.1600-0773.1978.tb02240.x Wang, X., Zhang, W. and Zhou, S. 2024. Multifaceted physiological and therapeutical impact of curcumin on hormone-related endocrine dysfunctions: a comprehensive review. Phytoth. Res. PTR. 38(7), 3307–3336; doi:10.1002/ptr.8208 Xiao, C. and Lai, D. 2025. Impact of oxidative stress induced by heavy metals on ovarian function. J. Appl. Toxicol. JAT. 45(1), 107–116; doi:10.1002/jat.4664 Yadav, A., Lomash, V., Samim, M. and Flora, S.J. 2012. Curcumin encapsulated in chitosan nanoparticles: a novel strategy for the treatment of arsenic toxicity. Chem. Biol. Interact. 199(1), 49–61; doi:10.1016/j.cbi.2012.05.011 Yallapu, M.M., Jaggi, M. and Chauhan, S.C. 2012. Curcumin nanoformulations: a future nanomedicine for cancer. Drug. Discovery. Today. 17(1-2), 71–80; doi:10.1016/j.drudis.2011.09.009 Yang, S.H., He, J.B., Yu, L.H., Li, L., Long, M., Liu, M.D. and Li, P. 2019. Protective role of curcumin in cadmium-induced testicular injury in mice by attenuating oxidative stress via Nrf2/ARE pathway. Environ. Sci. Pollut. Res. Int. 26(33), 34575–34583; doi:10.1007/s11356-019-06587-9 Zhang, J., Zheng, S., Wang, S., Liu, Q. and Xu, S. 2020. Cadmium-induced oxidative stress promotes apoptosis and necrosis through the regulation of the miR-216a-PI3K/AKT axis in common carp lymphocytes and antagonized by selenium. Chemosphere 258, 127341; doi:10.1016/j.chemosphere.2020.127341 Zhao, M., Ma, J., Li, M., Zhang, Y., Jiang, B., Zhao, X., Huai, C., Shen, L., Zhang, N., He, L. and Qin, S. 2021. Cytochrome P450 enzymes and drug metabolism in humans. Int. J. Mol. Sci. 22(23), 12808; doi:10.3390/ijms222312808 | ||
| How to Cite this Article |
| Pubmed Style Al-muhja MNA, Al-saaidi JAA. Nano-formulated curcumin restores hypothalamic–pituitary–ovarian axis disturbance in cadmium-induced female reproductive toxicity. Open Vet. J.. 2026; 16(5): 2619-2630. doi:10.5455/OVJ.2026.v16.i5.5 Web Style Al-muhja MNA, Al-saaidi JAA. Nano-formulated curcumin restores hypothalamic–pituitary–ovarian axis disturbance in cadmium-induced female reproductive toxicity. https://www.openveterinaryjournal.com/?mno=311008 [Access: June 26, 2026]. doi:10.5455/OVJ.2026.v16.i5.5 AMA (American Medical Association) Style Al-muhja MNA, Al-saaidi JAA. Nano-formulated curcumin restores hypothalamic–pituitary–ovarian axis disturbance in cadmium-induced female reproductive toxicity. Open Vet. J.. 2026; 16(5): 2619-2630. doi:10.5455/OVJ.2026.v16.i5.5 Vancouver/ICMJE Style Al-muhja MNA, Al-saaidi JAA. Nano-formulated curcumin restores hypothalamic–pituitary–ovarian axis disturbance in cadmium-induced female reproductive toxicity. Open Vet. J.. (2026), [cited June 26, 2026]; 16(5): 2619-2630. doi:10.5455/OVJ.2026.v16.i5.5 Harvard Style Al-muhja, M. N. A. & Al-saaidi, . J. A. A. (2026) Nano-formulated curcumin restores hypothalamic–pituitary–ovarian axis disturbance in cadmium-induced female reproductive toxicity. Open Vet. J., 16 (5), 2619-2630. doi:10.5455/OVJ.2026.v16.i5.5 Turabian Style Al-muhja, Mohammed Nabeel Abbas, and Jabbar Abbas Ahmed Al-saaidi. 2026. Nano-formulated curcumin restores hypothalamic–pituitary–ovarian axis disturbance in cadmium-induced female reproductive toxicity. Open Veterinary Journal, 16 (5), 2619-2630. doi:10.5455/OVJ.2026.v16.i5.5 Chicago Style Al-muhja, Mohammed Nabeel Abbas, and Jabbar Abbas Ahmed Al-saaidi. "Nano-formulated curcumin restores hypothalamic–pituitary–ovarian axis disturbance in cadmium-induced female reproductive toxicity." Open Veterinary Journal 16 (2026), 2619-2630. doi:10.5455/OVJ.2026.v16.i5.5 MLA (The Modern Language Association) Style Al-muhja, Mohammed Nabeel Abbas, and Jabbar Abbas Ahmed Al-saaidi. "Nano-formulated curcumin restores hypothalamic–pituitary–ovarian axis disturbance in cadmium-induced female reproductive toxicity." Open Veterinary Journal 16.5 (2026), 2619-2630. Print. doi:10.5455/OVJ.2026.v16.i5.5 APA (American Psychological Association) Style Al-muhja, M. N. A. & Al-saaidi, . J. A. A. (2026) Nano-formulated curcumin restores hypothalamic–pituitary–ovarian axis disturbance in cadmium-induced female reproductive toxicity. Open Veterinary Journal, 16 (5), 2619-2630. doi:10.5455/OVJ.2026.v16.i5.5 |

