




| Research Article | ||
Open Vet. J.. 2026; 16(5): 2659-2666 Open Veterinary Journal, (2026), Vol. 16(5): 2659-2666 Research Article Evaluation of immunological and gene expression responses induced by Galectin-3 and whole-sonicated Candida krusei antigen in rats experimentally infected with Candida kruseiHussein Ali Hussein* and Inam Bader FalihDepartment of Pathology and Poultry Diseases, College of Veterinary Medicine, University of Baghdad, Baghdad, Iraq *Corresponding Author: Hussein Ali Hussein. Department of Pathology and Poultry Diseases, College of Veterinary Medicine, University of Baghdad, Baghdad, Iraq. Email: hussein.ali2207p [at] covm.uobaghdad.edu.iq Submitted: 23/01/2026 Revised: XX/XX/XX Accepted: 08/03/2026 Published: XX/XX/XX © 2025 Open Veterinary Journal
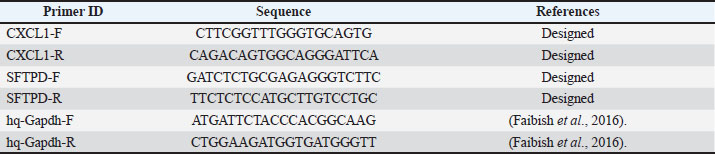
ABSTRACTBackground: Candida krusei is an emerging non-Candida albicans species increasingly associated with antifungal resistance and invasive systemic infections. Galectin-3 (GAL-3), a β-galactosidase-binding lectin, and the surfactant protein D gene are important components of innate antifungal immunity. The chemokine (C-X-C motif) ligand-1 gene plays a central role in neutrophil recruitment, whereas whole-sonicated C. krusei antigen may act as a broad fungal immunogen. Aim: To evaluate the immunological and pulmonary gene expression responses induced by GAL-3, whole-sonicated C. krusei antigen, and their combination in rats experimentally infected with C. krusei, via measurement of serum immunoglobulin G, interleukin-17, interleukin-23, and lung expression of Chemokine (C-X-C motif) ligand 1 (CXCL-1) and surfactant protein D genes. Methods: A total of 120 rats were randomly allocated into four experimental groups (n=30). Animals received GAL-3, whole-sonicated C. krusei antigen, and their combination, while the control group was subdivided into infected and healthy controls. Immunized groups were administered subcutaneously according to the experimental schedule. Systemic C. krusei infection was induced intraperitoneally, and samples were collected at 15 and 30 days post-challenge. Serum Immunoglobulin G, Interleukin -17 (IL-17), and Interleukin -23 (IL-23) levels were measured using ELISA. Pulmonary CXCL-1 and surfactant protein D gene expression were quantified using SYBR Green reverse transcription quantitative polymerase chain reaction and analyzed using the 2⁻ΔΔCt method. Results: Immunoglobulin G levels increased significantly in all treated groups compared with infected and healthy controls, with the highest levels at day 30. Whole-sonicated C. krusei antigen and mixed treatment groups showed significant increases in IL-17 and IL-23, indicating activation of Th17-mediated immune responses. Galectin-3 treatment produced upregulation of CXCL-1 and surfactant protein D genes, whereas the antigen-treated group showed moderate gene expression changes. The combined treatment group demonstrated reduced CXCL-1 induction but maintained a modest increase in surfactant protein D expression. Conclusion: Galectin-3 and whole-sonicated C. krusei antigen may enhance antifungal immune responses through partially distinct mechanisms. Galectin-3 modulates the CXCL-1 and surfactant protein D gene axis and activate innate immune pathways, whereas whole-sonicated antigen stimulates IL-23/IL-17-mediated responses and humoral immunity. These findings suggest that GAL-3 and fungal antigen immunization may represent support immunomodulatory approaches for controlling infections caused by non-C. albicans Keywords: Candida krusei, Galectin-3, Non-Candida albicans species, Surfactant protein D, Whole sonicated antigen. IntroductionGalectin-3 (GAL-3) is a multifunctional member of the galectin family characterized by its β-galactoside–binding activity and the presence of Carbohydrate Recognition Domains. It interacts with target molecules through carbohydrate-dependent and -independent mechanisms and is primarily localized in the cytoplasm under physiological conditions (Bouffette et al., 2023). Galectin-3 can be released into the extracellular environment during inflammation or tissue injury, where it functions as a diagnostic and prognostic biomarker in several pathological conditions. Increasing evidence also indicates that GAL-3 plays an important role in antifungal host defense, particularly by enhancing Candida species neutrophil phagocytosis (Wu et al., 2017; Verma et al., 2019). Non-Candida albicans species exhibit considerable phenotypic diversity, including variations in morphology, cell wall composition, and virulence factor expression. These differences influence fungal recognition by the host immune system and contribute to variable clinical outcomes. In this context, GAL-3 has emerged as an immunomodulatory molecule that may exert both protective and immunosuppressive effects depending on the infecting species and host immune status (Whibley et al., 2015). The Interleukin-23 (IL-23)/Interleukin-17 (IL-17) cytokine axis plays a central role in antifungal immunity. IL-17 and IL-23 are essential for neutrophil recruitment and the induction of antimicrobial peptides that contribute to controlling fungal proliferation, including infections caused by Candida krusei (Mengesha and Conti, 2017; Ivasheva et al., 2022). Chemokine (C-X-C motif) ligand 1 (CXCL-1) plays an important role in the recruitment and activation of neutrophils during fungal infection. Several studies have suggested that CXCL-1 expression may be influenced by upstream inflammatory cytokines, including IL-17, as part of a coordinated innate immune signaling pathway rather than a direct single-step mechanism (Lu et al., 2025). In addition to chemokine-mediated recruitment of immune cells, other innate immune molecules contribute to early antifungal defense. Surfactant protein D (SP-D) is a critical pattern-recognition molecule that binds to fungal cell wall components, promoting aggregation and enhancing opsonization, thereby facilitating neutrophil and macrophage phagocytosis. Furthermore, SP-D has been reported to interfere with fungal adhesion to host tissues and to reduce hyphal and pseudo-hyphal formation, supporting early mucosal and systemic defense against Candida species (Ordonez et al., 2019; Madan and Kishore, 2020). This study aimed to evaluate the immunological and molecular effects of GAL-3 and whole-sonicated C. krusei antigen, administered individually and in combination, on host defense against experimental C. krusei infection in rats by assessing cytokine responses (IL-17 and IL-23), Immunoglobulin G (IgG) levels ,and gene expression (CXCL-1 and Surfactant Protein D (SFTPD)). Materials and MethodsExperimental designA total of 120 rats were randomly allocated into 4 experimental groups. The GAL-3-treated animals group received subcutaneous administration of GAL-3 in two immunization doses on days 0 and 14. The whole-sonicated C. krusei antigen group received subcutaneous injections following the same immunization schedule. The combined treatment group received both GAL-3 and whole sonication C. krusei antigen concurrently administered on days 0 and 14. The control group was maintained under identical experimental conditions and further subdivided into a positive control subgroup subjected to infection and a negative control subgroup receiving sterile phosphate-buffered saline (PBS). All animals were challenged intraperitoneally with an infectious dose of C. krusei on day 29 from the start of the experiment. Blood and tissue samples were subsequently collected at 15 and 30 days post-challenge for cytokine level (IL-17 and IL-23) evaluation and gene expression analysis. Estimation of Galectin-3 (GAL-3) doseThe dose of GAL-3 was estimated based on the study of Verma et al. (2019) with slight modification. The mouse dose corresponded to approximately 1 mg/kg, and the equivalent rat dose was calculated as 0.5 mg/kg using body-surface-area allometric scaling (Km_mouse=3, Km_rat=6). For a 150-g rat, this equals 0.075 mg (75 µg). This calculated dose was administered twice (day 0 and day 14) as a prime-boost immunization schedule, resulting in a total administered dose of 150 µg per rat before challenge. Preparation of whole-sonicated Candida krusei antigenWhole-sonicated C. krusei antigen was prepared according to the method of Mohammed and Al-Samarraae (2021) with minor modifications. A dense culture of C. krusei was grown on Sabouraud dextrose agar at 37°C for 48 hours. Fungal cells were harvested using (PBS, pH 7.2), washed, and fixed in 0.5% formalinized PBS. Subsequently, the suspension was sonicated using a Soniprep 150 ultrasonic device (20 kHz) under cold conditions, followed by centrifugation and filtration through a 0.45 µm membrane filter. Sterility and complete cell disruption were confirmed by Sabouraud Dextrose Agar culture. Protein concentration was determined using the Lowry method (Lowry et al., 1951), and a protein concentration of 5 mg was used as the immunization dose. Determination of Candida krusei infectious doseExperimental C. krusei infection was induced by intraperitoneal injection (Anaissie et al., 1993), with minor modifications. A preliminary pilot study was conducted using a small group of rats (n=10) to determine the optimal infectious dose capable of producing consistent infection without causing acute mortality. Based on the findings of the pilot study, an inoculum of 1 × 10⁸ CFU per animal was selected as the infectious challenge dose for the main experiment. Animals were monitored daily for clinical signs of infection and survival for 7 days after infection. Tissue samplesLung tissue samples were collected from all groups, including the whole sonicated antigen group, GAL-3 group, and mix group (whole-sonicated C. Krusei antigen + GAL-3), as well as from healthy negative and positive infected control groups at 15 and 30 days post-challenge infection. Each sample was placed in 600 μl of GENEzol™ reagent in an Eppendorf tube and stored for further analysis. IL-17, IL-23 and IgG quantificationSerum IL-17, IL-23, and IgG concentrations were measured using rat Enzyme-Linked Immunosorbent Assay kits (GenoChem, Spain) according to the manufacturer’s protocol. Blood samples were collected from all groups at three time points (Parasuraman et al., 2010): 29 days before infection and 15 and 30 days post-challenge. Serum was separated by centrifugation at 3,000 rpm for 15 minutes and stored at −20°C until analysis. The optical density was measured at 450 nm, and the standard curve was used to calculate cytokine and IgG levels. Statistical analysis was performed using two-way analysis of variance, with significance set at p ≤ 0.05. Ribonucleic Acid (RNA) extraction and cDNA synthesisTotal RNA was extracted from the lung tissue of all groups by using the GENEzol™ Reagent Kit (Geneaid, Korea) according to the manufacturer’s protocol. cDNA was synthesized from 5 μl of total RNA using the Transcript 1 gDNA Removal and cDNA Synthesis Super Mix Kit. Protocol (Al-Khafaji et al., 2025). Relative quantitative polymerase chain reaction (PCR) conditions and analysisQuantitative Polymerase Chain Reaction (qPCR) was performed using the TransStart Top Green qPCR SuperMix kit to monitor dsDNA synthesis. Each reaction was performed in a final volume of 20 μl, containing 10 μl of Green Master Mix reagent, 1 μl of each primer, 3 μl of cDNA, and 5 μl of nuclease-free water. The thermal cycling protocol consisted of an initial denaturation at 94°C for 30 seconds, followed by 40 cycles of denaturation at 94°C for 5 seconds, annealing at 58°C for 15 seconds, and extension at 72°C for 20 seconds. A melting curve analysis from 65°C to 95°C was performed at the end of the run to verify the specificity of the amplification. Primer selection and preparationAll primers for the target and housekeeping genes were synthesized by Macrogen® (Korea) according to the primer designs prepared at Nabu Corporation (Baghdad). The primer sequences and characteristics are listed in Table 1. Table 1. Primers sequences designed for the target gene and the housekeeping gene.
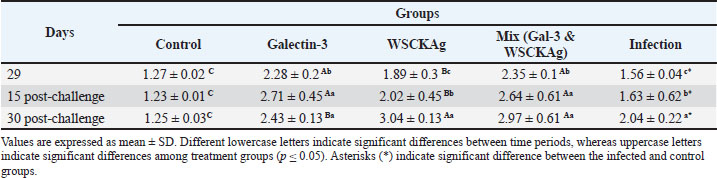
Calculation of the CXCL-1 and SFTPD genes expressionGene expression levels were normalized to the housekeeping gene (GAPDH) and calculated using the 2−ΔΔCt method (Schmittgen and Livak, 2008) ΔCt=Ct (target gene)—Ct (GAPDH), ΔΔCt=ΔCt(sample)—average ΔCt (control group), Fold change=2⁻ΔΔCt. Fold-change values >1 indicated upregulation, whereas values < 1 indicated downregulation. Ethical approvalThis study was approved by the Ethics Committee of the College of Veterinary Medicine, University of Baghdad, Baghdad, Iraq (Code No. 39/PG, approved on January 7, 2026). ResultsMeasurement of IgG and cytokine levels (IL-17 and IL-23)The results of the present study showed a significant increase in IgG levels at days 15 post-challenge infection with groups for GAL-3, whole-sonicated C. krusei Antigen (WSCKAg), and mixed (Gal-3 and WSCKAg), respectively, when compared with baseline values before infection (day 29) for (GAL-3), sonicated (WSCKAg), and mixed group (Gal-3 and WSCKAg), respectively, and with control group, and continued to increase to the end of study at day 30, respectively, as shown in Table 2 Table 2. Comparison of serum IgG levels among experimental groups at different time points.
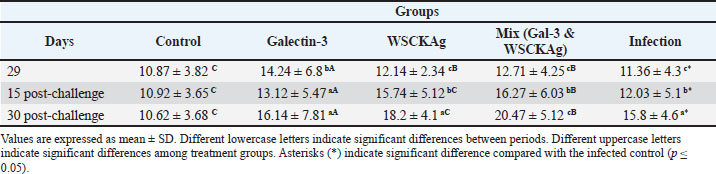
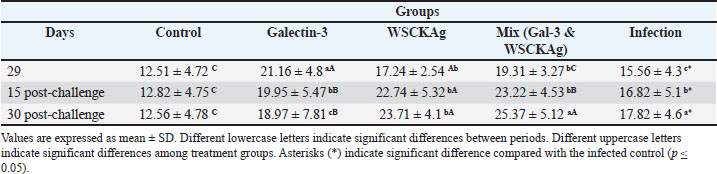
As shown in Table 3, IL-17 and cytokine levels demonstrated a progressive increase in the WSCKAg and mixed groups at both 15 and 30 days post-challenge compared with day 29 and the control group. Galectin-3 treatment also showed an increase at day 30 compared with day 15, suggesting enhanced NAI responses during the later phase of infection. Similarly, IL-23 levels followed a comparable pattern, with significant increases observed in the WSCKAg and mixed groups at both post-challenge time points (Table 4). Although GAL-3 showed moderately elevated IL-23 levels, the mixed treatment group exhibited the highest concentrations at day 30 post-challenge, indicating a stronger activation of the IL-23/IL-17 immune axis. Table 3. Comparison of serum IL-17 levels among experimental groups at different time points.
Table 4. Comparison of serum IL-23 levels among experimental groups at different time points.
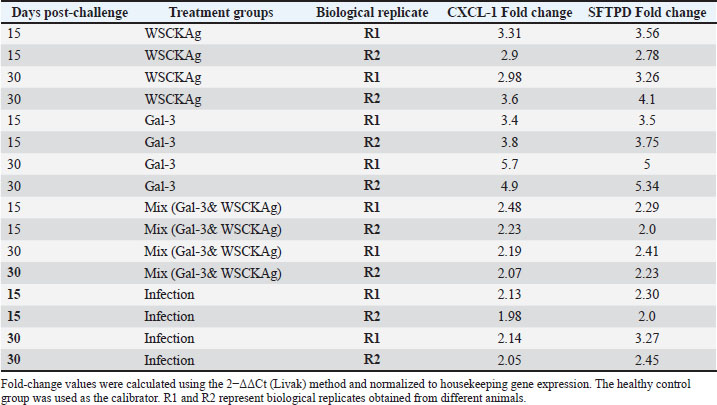
Expression of CXCL-1 and SFTPD in major infected organs following galectin-3 treatment and post-challenge injectionThe present findings are shown in Table 5 and Figure 1. Upregulation of the expression of CXCL-1 and SFTPD gene was observed. In the WSCKAg group, CXCL-1 expression showed a moderate increase, accompanied by a comparable elevation of SFTPD relative to the control. Galectin-3 treatment alone induced the highest upregulation of both genes, particularly on day 30. In contrast, the mixed group (Gal-3 + WSCKAg) exhibited only minimal changes in CXCL-1 and a modest upregulation in SFTPD. The infection group exhibited intermediate fold changes for both CXCL-1 and SFTPD. Table 5. Fold-change expression of CXCL1 and SFTPD genes in experimental groups at 15 and 30 days post-infection.
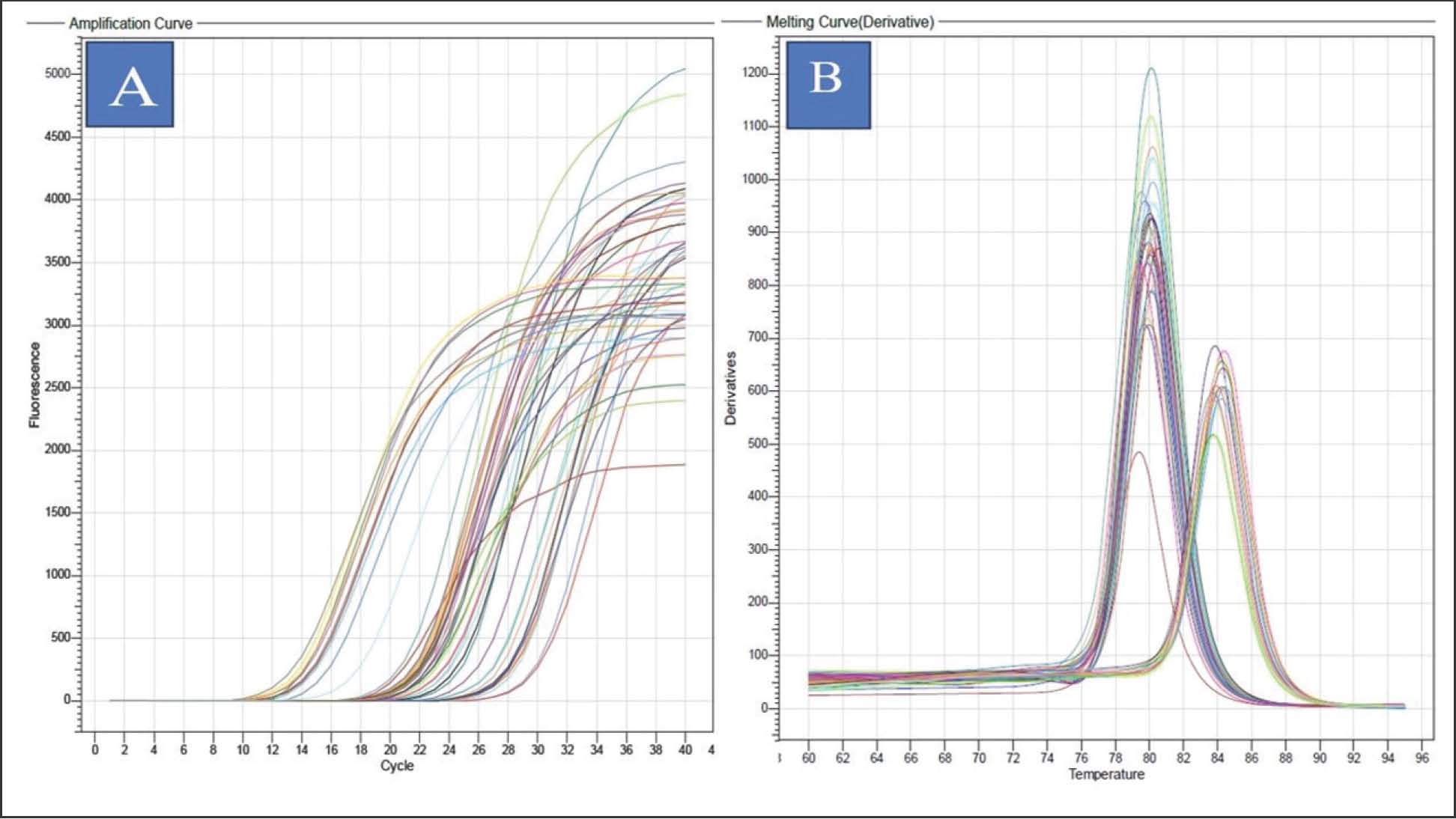
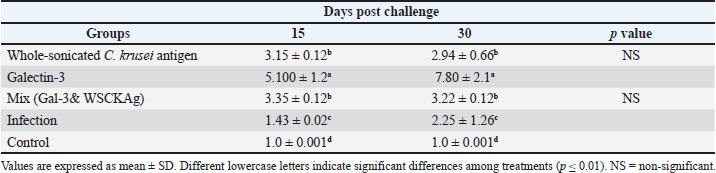
Fig. 1. (A): Amplification curves of CXCL1, SFTPD, and the housekeeping gene across the different experimental groups. (B): Efficient exponential amplification and reproducible Ct values among replicates. The result in Table 6 shows the comparison of ΔCt values (mean ± SD) for CXCL-1 and SFTPD among the different experimental groups at 15 and 30 days. On day 15, the GAL-3-treated rats exhibited the highest gene expression, which was significantly higher than that of the WSCKAg and Gal-3 + WSCKAg mix groups, the infection group, and the control group at p ≤ 0.01. A similar pattern was observed at day 30, where GAL-3 still showed the greatest upregulation, WSCKAg and the mix group showed intermediate values, and the infection group showed a modest increase compared with the control. Table 6. Comparison of ΔCt values (mean ± SD) for CXCL-1 and SFTPD genes among experimental groups.
DiscussionThe present findings may contribute to enhanced antifungal immune responses following immunization with whole-sonicated C. krusei antigen (WSCKAg), Gal-3, and their combined administration after experimental infection. As shown in Table 2, a significant elevation in IgG levels was observed in the treated groups compared with the infected controls. Similar findings have been reported in studies demonstrating that sonicated fungal antigens stimulate antibody production through enhanced antigen presentation, B-cell activation, and antibody-mediated immune defense (Nada et al., 2018; Alrudah et al., 2024). Galectin-3 is recognized as an immunomodulatory lectin involved in both innate and adaptive immune regulation. Gal-3 enhances antigen uptake and presentation by macrophages and dendritic cells, thereby promoting B-lymphocyte activation and antibody production (Díaz-Alvarez and Ortega, 2017; Sciacchitano et al., 2018). These mechanisms may explain the increased IgG levels observed in the Gal-3-treated group in the present study. Rezende et al. (2021a) reported that GAL-3 deficiency impairs IgG-mediated antifungal immunity through altered Fcγ receptor-dependent functions, including opsonization and phagocytosis. The sustained increase observed in the mixed-treatment group suggests a potential synergistic interaction between WSCKAg immunization and GAL-3-mediated immune modulation. Cytokine analysis further highlighted the importance of the IL-23/IL-17 axis in antifungal defense. As demonstrated in Tables 3 and 4, IL-23 regulates Th17 cell differentiation and stabilization, whereas IL-17 promotes neutrophil recruitment and antimicrobial peptide production essential for fungal clearance (Mengesha and Conti, 2017; Iwasawa et al., 2022). Similar cytokine elevations following fungal antigen immunization have been described previously (Kadhim and Faleh, 2021). In addition, Sawada et al. (2021) indicated that IL-17 signaling is essential for antifungal immunity in controlling Candida spp. infections. In addition, the present findings regarding the effects of GAL-3 on cytokines are in agreement with those reported by Rezende et al. (2021a), who demonstrated that GAL-3 deficiency reduces IL-23 expression and subsequently impairs IL-17 responses. The higher IL-23 levels observed on day 15 compared with IL-17 may reflect its upstream regulatory role in stabilizing Th17 differentiation and promoting IL-17 production, which contributes to neutrophil recruitment and antimicrobial defense against extracellular fungal pathogens (Dillon et al., 2025). The gene expression findings further supported these immunological observations. Galectin-3 and WSCKAg administration resulted in upregulation of CXCL-1 and SFTPD genes (Tables 5 and 6) and (Figs. 1 and 2),, respectively. CXCL-1 is a potent neutrophil chemoattractant responsible for early recruitment of immune cells during fungal invasion (Radam and Faleh, 2015; Yanhua et al., 2025). Neutrophils represent a central component of innate antifungal immunity through phagocytosis and neutrophil extracellular trap formation. These findings are consistent with those of Snarr et al. (2020), who demonstrated reduced neutrophil recruitment, which led to increased susceptibility to fungal infection.
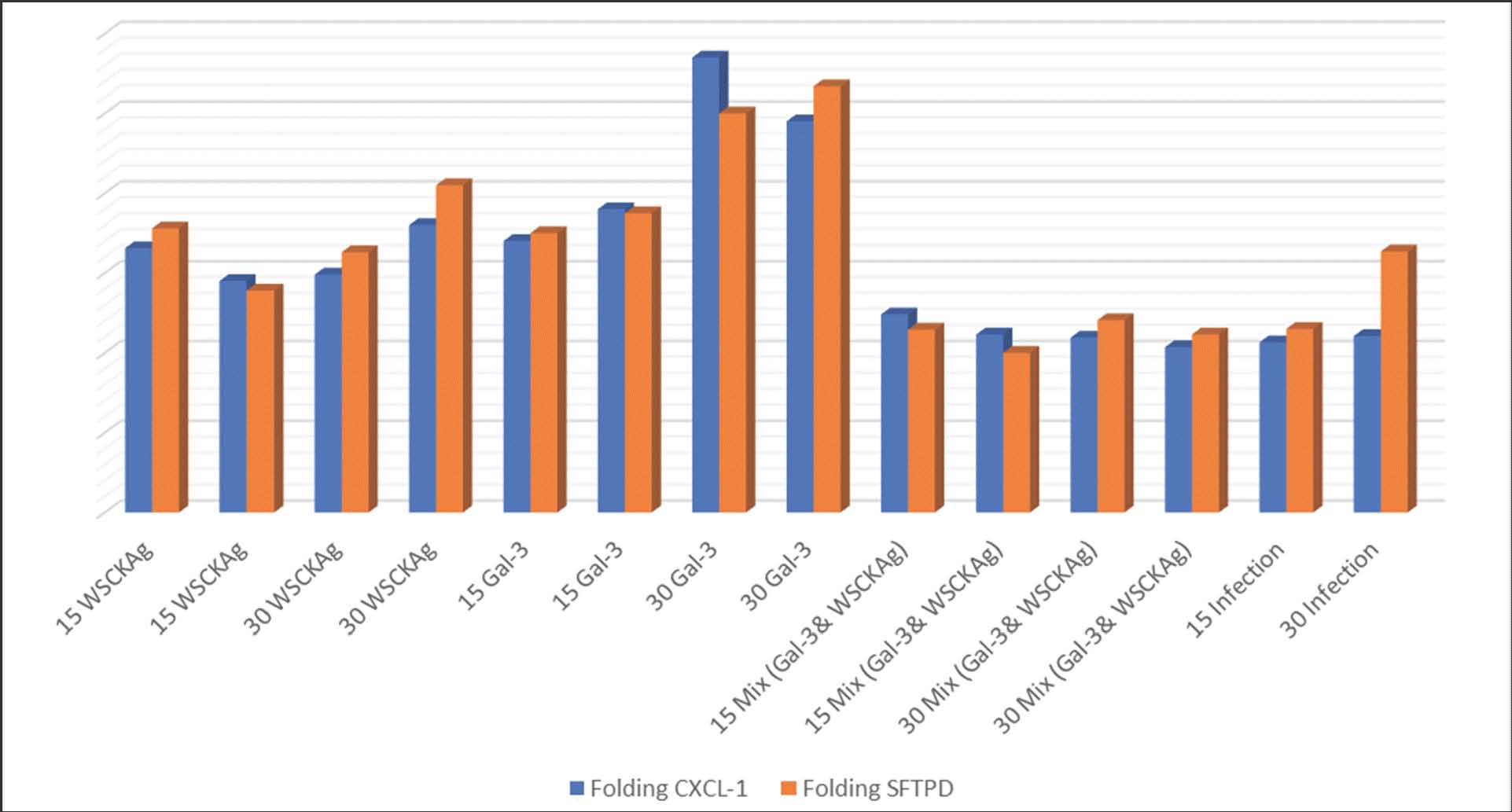
Fig. 2. Relative expression levels (fold change) of CXCL-1 and SFTPD genes in experimental groups treated with WSCKAg, Galectin-3, their combination (MIX), and infection at 15- and 30-days post-challenge infection. In addition, the SFTPD gene encodes SP-D, a pattern-recognition molecule that binds fungal cell-wall carbohydrates, such as mannans and glucans, promoting opsonization and phagocytic clearance (Madan and Kishore, 2020). Therefore, increased SFTPD expression suggests activation of SP-D-mediated antifungal defense mechanisms. Similarly, the present findings align with previous studies (Rezende et al., 2021a, b) that reported the dysregulation of innate immune genes, including CXCL-1 and SFTPD, following GAL-3 deletion in experimental murine models. WSCKAg immunization also promoted CXCL-1 expression, which is consistent with the results of Bai et al. (2020), who reported enhanced chemokine production following fungal antigen exposure. Furthermore, Ordoñez et al. (2019) demonstrated the fungistatic and fungicidal activity of SP-D through direct binding to fungal surfaces and enhancement of phagocytic clearance. ConclusionGalectin-3 and whole-sonicated C. krusei antigen may enhance antifungal immunity through partially distinct immunological mechanisms. Galectin-3 appears to be associated with modulation of the CXCL-1 and surfactant protein D gene axis and activation of innate effector pathways, whereas whole-sonicated C. krusei antigen may be associated with increased IL-23/IL-17 responses and humoral immune activation. The coordinated regulation of CXCL-1 and surfactant protein D suggests a potential role for these molecules as important components of host defense against C. krusei. Collectively, these findings indicate that GAL-3 and whole-sonicated C. krusei antigen may represent promising candidates for further investigation as immunomodulatory strategies in non-albicans Candida infections. AcknowledgmentThe authors gratefully acknowledge the College of Veterinary Medicine, University of Baghdad, for providing laboratory facilities and technical support during this study. Conflicts of interestThe authors declare no conflicts of interest. FundingNone. Authors' contributionsAll authors contributed equally to this work and approved the final version of the manuscript. Data availabilityThe datasets generated and analyzed during the current study are available upon reasonable request from the corresponding author. ReferencesAlrudah, J.H.A., Abbas, N.F., Hadi, F.A. and Kaab, H.T. 2024. Immunization protocol for whole cell fraction antigen extracted from Candida albicans isolated from pigeon with candidiasis. J. Anim. Health Prod. 12(S1), 107–112; doi:10.17582/journal.jahp/2024/12.s1.107 Anaissie, E., Hachem, R., K-Tin-U, C., Stephens, L.C. and Bodey, G.P. 1993. Experimental hematogenous candidiasis caused by Candida krusei and Candida albicans: species differences in pathogenicity. InfectImmunity 61(4), 1268–1271; doi:10.1128/iai.61.4.1268-1271.1993 Bai, W., Wang, Q., Deng, Z., Li, T., Xiao, H. and Wu, Z. 2020. TRAF1 suppresses antifungal immunity through CXCL1-mediated neutrophil recruitment during Candida albicans intradermal infection. Cell. Communication. Signaling. 18(1), 30; doi:10.1186/s12964-020-00532-x Bouffette, S., Botez, I. and De Ceuninck, F. 2023. Targeting galectin-3 in inflammatory and fibrotic diseases. Trends. Pharmacol. Sci. 44(8), 519–531; doi:10.1016/j.tips.2023.06.001 Díaz-Alvarez, L. and Ortega, E. 2017. The many roles of galectin-3, a multifaceted molecule, in innate immune responses against pathogens. Mediators Inflammation 2017, 9247574; doi:10.1155/2017/9247574 Dillon, J.T., Hickey, M.T., Saul-Mcbeth, J., Glanz, T., Daboul, Y., Byreddy, Y.R., Hohl, R., Kratch, J.M., Thakker, J.A., Gaffen, S.L., Gibbs, J. and Conti, H.R. 2025. Synergistic exacerbation of oral mucositis caused by IL-23 deficiency and oral Candida albicans exposure. mBio 16(10), e01992-25; doi:10.1128/mbio.01992-25 Faibish, D., Suzuki, M. and Bartlett, J.D. 2016. Appropriate real-time PCR reference genes for fluoride treatment studies performed in vitro or in vivo. Arch. Oral Biol. 62, 33–42; doi: 10.1016/j.archoralbio.2015.11.004 Hiba Muneer Abd-Alhassan Al-Khafaji., Rehab M. Khadum., Sara Mohammed Ouda. and Maryam Qasim Mohammed. 2025. Inverse correlation of miR-196a and HOXB13 expression in Iraqi patients with prostate cancer. Cellular Mol. Biol. 71(9), 63–70; doi:10.14715/cmb/2025.71.9.8 Iwasawa, M.T., Miyachi, H., Wakabayashi, S., Sugihira, T., Aoyama, R., Nakagawa, S., Katayama, Y., Yoneyama, M., Hara, H., Iwakura, Y., Matsumoto, M., Inohara, N., Koguchi-Yoshioka, H., Fujimoto, M., Núñez, G., Matsue, H., Nakamura, Y. and Saijo, S. 2022. Epidermal clearance of Candida albicans is mediated by IL-17 but independent of fungal innate immune receptors. Int. Immunol. 34(8), 409–420; doi:10.1093/intimm/dxac019 Kadhim, S.O. and Faleh, I.B. 2021. Pathological study of genitourinary invasion by Aspergillus flavus in male rats. Iraqi. J. Vet. Sci. 35 (Supplement I-III), 87–94; doi:10.33899/ijvs.2021.131580.1974 Lowry, O., Rosebrough, N., Farr, A.L. and Randall, R. 1951. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 193(1), 265–275; doi:10.1016/S0021-9258(19)52451-6 Lu, J., Dong, Q., Zhang, R., Wu, W. and Yang, H. 2025. CXCL-1 and CXCL-2: key regulators of host defense against Phialophora verrucosa. J. Inflammation. Res. 6319, 6319–6329; doi:10.2147/JIR.S518653 Madan, T. and Kishore, U. 2020. Surfactant protein D recognizes multiple fungal ligands: a key step to initiate and intensify the antifungal host defense. Front. Cellular InfectMicrobiol. 10, 229; doi:10.3389/fcimb.2020.00229 Mengesha, B. and Conti, H. 2017. The role of IL-17 in protection against mucosal Candida infections. J. Fungi. 3(4), 52; doi:10.3390/jof3040052 Mohammed, R.J. and Al-Samarraae, I.A.A. 2021. Investigating the effect of three antigens of Citrobacter freundii on rabbit’s immune response. Iraqi. J. Vet. Med. 45(1), 56–62; doi:10.30539/ijvm.v45i1.1043 Nada, F.A., Shani, W.S. and Najim, I.M. 2018. Evaluation of immunization protocol in mice injected with whole cell fraction antigen of Candida albicans isolated from vaginal infections. Scientific. J. Med. Res. 2, 42–46; doi:10.37623/SJMR.2018.2509 Ordonez, S.R., Van Eijk, M., Escobar Salazar, N., De Cock, H., Veldhuizen, E.J.A. and Haagsman, H.P. 2019. Antifungal activities of surfactant protein D in an environment closely mimicking the lung lining. Mol. Immunol. 105, 260–269; doi:10.1016/j.molimm.2018.12.003 Parasuraman, S., Raveendran, R. and Kesavan, R. 2010. Blood sample collection in small laboratory animals. J. Pharmacol. Pharmacother. 1(2), 87–93; doi: 10.4103/0976-500X.72350 Radam, E.B. and Faleh, S.A. 2015. Pathological effect of experimental Candida albicans infection in mice immunized with whole killed C. albicans lyophilized antigen. In: Proceedings of the 3rd International Scientific Conference of Genetic and Environment, Baghdad, Iraq, 2015, pp 555–571. Rezende, C.P., Brito, P.K.M.O., Da Silva, T.A., Pessoni, A.M., Ramalho, L.N.Z. and Almeida, F. 2021. Influence of galectin-3 on the innate immune response during experimental cryptococcosis. J. Fungi 7(6), 492; doi:10.3390/jof7060492 Rezende, C.P., Martins Oliveira Brito, P.K., Pessoni, A.M., Da Silva, T.A., Goldman, G.H. and Almeida, F. 2021. Altered expression of genes related to innate antifungal immunity in the absence of galectin-3. Virulence 12(1), 981–988; doi:10.1080/21505594.2021.1903212 S, P., R, R. and R, K. 2010. Blood sample collection in small laboratory animals. J. Pharmacol. Pharmacotherapeutics. 1(2), 87–93; doi:10.4103/0976-500X.72350 Sawada, Y., Setoyama, A., Sakuragi, Y., Saito-Sasaki, N., Yoshioka, H. and Nakamura, M. 2021. The role of IL-17-producing cells in cutaneous fungal infections. Int. J. Mol. Sci. 22(11), 5794; doi:10.3390/ijms22115794 Schmittgen, T.D. and Livak, K.J. 2008. Analyzing real-time PCR data by the comparative CT method. Nat. Protocols 3(6), 1101–1108; doi:10.1038/nprot.2008.73 Sciacchitano, S., Lavra, L., Morgante, A., Ulivieri, A., Magi, F., De Francesco, G., Bellotti, C., Salehi, L. and Ricci, A. 2018. Galectin-3: one molecule for an alphabet of diseases, from A to Z. Int. J. Mol. Sci. 19(2), 379; doi:10.3390/ijms19020379 Snarr, B.D., St-Pierre, G., Ralph, B., Lehoux, M., Sato, Y., Rancourt, A., Takazono, T., Baistrocchi, S.R., Corsini, R., Cheng, M.P., Sugrue, M., Baden, L.R., Izumikawa, K., Mukae, H., Wingard, J.R., King, I.L., Divangahi, M., Satoh, M.S., Yipp, B.G., Sato, S. and Sheppard, D.C. 2020. Galectin-3 enhances neutrophil motility and extravasation into the airways during Aspergillus fumigatus infection. PLos Pathogens 16(8), e1008741; doi:10.1371/journal.ppat.1008741 Verma, P., Laforce-Nesbitt, S.S., Tucker, R., Mao, Q., De Paepe, M.E. and Bliss, J.M. 2019. Galectin-3 expression and effect of supplementation in neonatal mice with disseminated Candida albicans infection. Pediatric. Res. 85(4), 527–532; doi:10.1038/s41390-019-0279-x Whibley, N. and Gaffen, S.L. 2015. Beyond Candida albicans: mechanisms of immunity to non-albicans Candida species. Cytokine 76(1), 42–52; doi:10.1016/j.cyto.2015.07.025 Wu, S.Y., Huang, J.H., Chen, W.Y., Chan, Y.C., Lin, C.H., Chen, Y.C., Liu, F.T. and Wu-Hsieh, B.A. 2017. Cell-intrinsic galectin-3 attenuates neutrophil ROS-dependent killing of Candida by modulating CR3 downstream Syk activation. Front. Immunol. 8, 48; doi:10.3389/fimmu.2017.00048 Yanhua, R., Renyi, C., Li, W., Yu, W. and Chen, C. 2025. Molecular mechanisms of neutrophil regulatory network in anti-Candida infection. Front. Immunol. 16, 1716645; doi: 10.3389/fimmu.2025.1716645 | ||
| How to Cite this Article |
| Pubmed Style Hussein HA, Falih IB. Evaluation of immunological and gene expression responses induced by Galectin-3 and whole-sonicated Candida krusei antigen in rats experimentally infected with Candida krusei. Open Vet. J.. 2026; 16(5): 2659-2666. doi:10.5455/OVJ.2026.v16.i5.8 Web Style Hussein HA, Falih IB. Evaluation of immunological and gene expression responses induced by Galectin-3 and whole-sonicated Candida krusei antigen in rats experimentally infected with Candida krusei. https://www.openveterinaryjournal.com/?mno=307893 [Access: June 26, 2026]. doi:10.5455/OVJ.2026.v16.i5.8 AMA (American Medical Association) Style Hussein HA, Falih IB. Evaluation of immunological and gene expression responses induced by Galectin-3 and whole-sonicated Candida krusei antigen in rats experimentally infected with Candida krusei. Open Vet. J.. 2026; 16(5): 2659-2666. doi:10.5455/OVJ.2026.v16.i5.8 Vancouver/ICMJE Style Hussein HA, Falih IB. Evaluation of immunological and gene expression responses induced by Galectin-3 and whole-sonicated Candida krusei antigen in rats experimentally infected with Candida krusei. Open Vet. J.. (2026), [cited June 26, 2026]; 16(5): 2659-2666. doi:10.5455/OVJ.2026.v16.i5.8 Harvard Style Hussein, H. A. & Falih, . I. B. (2026) Evaluation of immunological and gene expression responses induced by Galectin-3 and whole-sonicated Candida krusei antigen in rats experimentally infected with Candida krusei. Open Vet. J., 16 (5), 2659-2666. doi:10.5455/OVJ.2026.v16.i5.8 Turabian Style Hussein, Hussein Ali, and Inam Bader Falih. 2026. Evaluation of immunological and gene expression responses induced by Galectin-3 and whole-sonicated Candida krusei antigen in rats experimentally infected with Candida krusei. Open Veterinary Journal, 16 (5), 2659-2666. doi:10.5455/OVJ.2026.v16.i5.8 Chicago Style Hussein, Hussein Ali, and Inam Bader Falih. "Evaluation of immunological and gene expression responses induced by Galectin-3 and whole-sonicated Candida krusei antigen in rats experimentally infected with Candida krusei." Open Veterinary Journal 16 (2026), 2659-2666. doi:10.5455/OVJ.2026.v16.i5.8 MLA (The Modern Language Association) Style Hussein, Hussein Ali, and Inam Bader Falih. "Evaluation of immunological and gene expression responses induced by Galectin-3 and whole-sonicated Candida krusei antigen in rats experimentally infected with Candida krusei." Open Veterinary Journal 16.5 (2026), 2659-2666. Print. doi:10.5455/OVJ.2026.v16.i5.8 APA (American Psychological Association) Style Hussein, H. A. & Falih, . I. B. (2026) Evaluation of immunological and gene expression responses induced by Galectin-3 and whole-sonicated Candida krusei antigen in rats experimentally infected with Candida krusei. Open Veterinary Journal, 16 (5), 2659-2666. doi:10.5455/OVJ.2026.v16.i5.8 |

