




| Research Article | ||
Open Vet. J.. 2026; 16(5): 2735-2745 Open Veterinary Journal, (2026), Vol. 16(5): 2735-2745 Research Article Optimizing sperm sexing in Bali cattle: Effects of separation time and bull age on BSA gradient efficiencyYendraliza Yendraliza1*, Muhamad Rodiallah1 and Jaswandi Jaswandi21Department of Animal Science, Faculty of Agriculture and Animal Science, Universitas Islam Sultan Syarif Kasim Riau, Pekanbaru, 28293, Riau, Indonesia 2Department of Animal Production Technology, Faculty of Animal Science, Universitas Andalas, Padang, West Sumatera, Indonesia *Corresponding Author: Yendraliza Yendraliza. Department of Animal Science, Faculty of Agriculture and Animal Science, Universitas Islam Sultan Syarif Kasim Riau, Pekanbaru, 28293, Riau, Indonesia. Email: yendraliza [at] uin-suska.ac.id Submitted: 01/12/2025 Revised: 15/03/2026 Accepted: 25/03/2026 Published: 31/05/2026 © 2025 Open Veterinary Journal
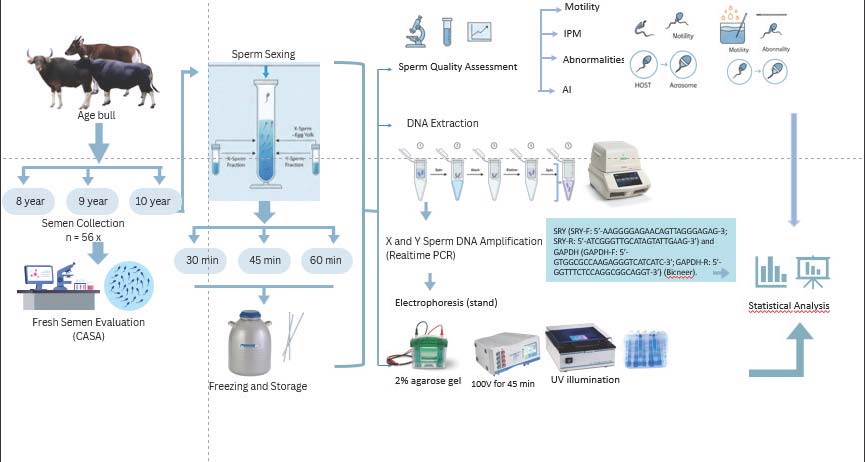
ABSTRACTBackground: Sperm sexing is an important biotechnology for increasing reproductive efficiency by aligning offspring sex with production goals. For Bali cattle (Bos javanicus), a native Indonesian breed with high genetic and cultural value, sexed semen also contributes to germplasm conservation and sustainable breeding programs. Bovine serum albumin (BSA) gradient separation offers a practical and affordable alternative to flow cytometry; however, separation time and donor bull age may influence the efficiency of this method. Aim: This study aimed to evaluate the effect of sperm separation time using the BSA method on sperm quality in bulls of different ages, before and after cryopreservation, confirmed by SRY–glyceraldehyde-3-phosphate dehydrogenase (GAPDH) amplification. Methods: Fifty-six ejaculates were collected from Bali bulls aged 8, 9, and 10 years. Samples were subjected to a factorial randomized block design with sperm separation times of 30, 45, and 60 minutes using 5% and 10% BSA gradients. The X- and Y-bearing fractions were verified through molecular amplification of the SRY and GAPDH genes. Semen quality parameters, including motility, viability, abnormality, plasma membrane integrity (IPM), and acrosome integrity, were evaluated before and after cryopreservation. Results: The separation time significantly influenced both molecular detection and post-thaw semen quality. A 30-minute separation period yielded the clearest discrimination of X and Y sperm and maintained higher post-thaw functional integrity than the 45- and 60-minute treatments. Among bull age groups, 9-year-old sires produced the highest quality semen across most parameters, particularly in terms of motility, viability, IPM, and acrosome integrity. Conclusion: Optimal conditions for BSA-based sperm sexing in Bali cattle are achieved with a 30-minute separation period using semen from 9-year-old bulls. These benchmarks provide a scientific basis for improving the efficiency of sexed semen production and support the sustainable use of Bali cattle in tropical breeding systems. Keywords: Bali cattle, Bovine serum albumin, Bull age, Cryopreservation, Sperm. IntroductionBali cattle (Bos javanicus) are one of Indonesia’s local cattle breeds that have been designated as national germplasm and have high economic, genetic, and cultural value (Agus and Mastuti Widi, 2018). The Bali cattle population was recorded at over 1.02 millions head in 2023, contributing significantly to the meat supply, food security, and income of small-scale farmers in eastern Indonesia (BPS, 2024). In addition to their excellent adaptation to the tropical climate, Bali cattle also have high reproductive efficiency, making them important candidates in local livestock breeding and conservation programs (Purwantara et al., 2012). The application of sperm sexing biotechnology has become a key strategy for improving production efficiency and accelerating genetic improvement in domestic livestock. This technology allows for sex selection, thereby enabling the targeted achievement of production goals, such as producing more females to increase the breeding population or more males for meat production (Garner and Seidel, 2008). Sexed semen has been shown to increase the economic value of the livestock industry by increasing genetic gain rates, improving management efficiency, and reducing the number of births that do not meet the intended goal (Butler et al., 2014). Biotechnology for sperm sexing has become a key strategy for improving production efficiency and accelerating genetic improvement in domestic livestock. Bovine serum albumin (BSA)-based sperm separation is a more economical and practical alternative in rural areas due to limited funds and the need for sophisticated equipment. The BSA technique utilizes the differences in density and composition of the X and Y spermatozoa membranes, allowing fractionation on an albumin gradient (Yendraliza et al., 2025). The effectiveness of the BSA method is heavily influenced by technical factors, such as separation duration and semen quality, which are closely related to the age of the donor stallion. Inappropriate separation duration can cause osmotic stress and decrease cell viability (Oldenhof et al., 2013). Meanwhile, the semen quality of bulls is influenced by age, where bulls that are too young or too old tend to have higher membrane damage, lower motility, and increased sensitivity to the freezing process (Warman et al., 2025). Successful sperm separation can be achieved through molecular testing using a combination of SRY–glyceraldehyde-3-phosphate dehydrogenase (GAPDH) with high accuracy and in a short time compared with in vivo testing (Mundana et al., 2023). sex determining region on Y (SRY) is a Y-chromosome–specific marker for identifying Y-bearing spermatozoa, whereas GAPDH serves as a housekeeping gene and internal control. Together, they enable accurate molecular verification of X- and Y-enriched sperm fractions after BSA-based separation. In Bali cattle, there are still very few scientific studies evaluating the effect of sperm separation time using the BSA method, the age of the bull, and the accuracy of separation based on molecular verification. These data gaps hinder the production of high-quality sexed semen to support national germplasm conservation and productivity improvement. Therefore, this study aimed to evaluate the effect of sperm separation time using the BSA method on sperm quality in bulls of different ages, before and after cryopreservation, as confirmed by SRY–GAPDH amplification. These findings are expected to provide applicable parameters for more efficient sexed semen production in tropical conditions and support the improvement of genetic conservation and productivity in Bali cattle. Materials and MethodsResearch locationThis study was conducted from September to October 2024 at the Regional Artificial Insemination Center (Balai Inseminasi Buatan Daerah), Tenayan Raya, Pekanbaru City, Riau Province, Indonesia. Animal handlingSemen samples were collected from Bali bulls aged 8, 9, and 10 years with body weights ranging from 512 to 609 kg. The bulls were housed individually and maintained under uniform management and feeding conditions. They exercised daily for 30 minutes by walking outside their stalls. Each animal received 60-kg Elephant Grass and 6-kg concentrate per day, divided into two feedings. Semen collection and processingSemen was collected once a week for 9 consecutive weeks using an artificial vagina (IMV, France) maintained at 42°C. Only ejaculates with a minimum motility of >70% and a sperm concentration of 800 × 10⁶/ml were used for further analysis. Each ejaculate was immediately evaluated for volume (ml), color, concentration (10⁶ sperm/ml), motility (%), viability (%), and abnormality (%) using a phase-contrast microscope. Experimental design and sperm separation using BSA columnsFigure 1 shows the overall experimental workflow, from semen collection to molecular verification. This study used a 2 × 3 factorial design in an randomized complete block design (RCBD). The first factor was the separation time during the incubation of the BSA column (30, 45, and 60 minutes). The second factor was bull age (8, 9, and 10 years). This age range was chosen to minimize the variability associated with sexual immaturity. The ejaculates from each bull served as the experimental units. Each ejaculate was independently processed through BSA separation, cryopreservation, thawing, and laboratory evaluation. A total of 56 ejaculates were collected over 9 consecutive weeks. Weekly semen collection served as a blocking factor to control temporal variation associated with collection time, environmental fluctuations, and handling differences.
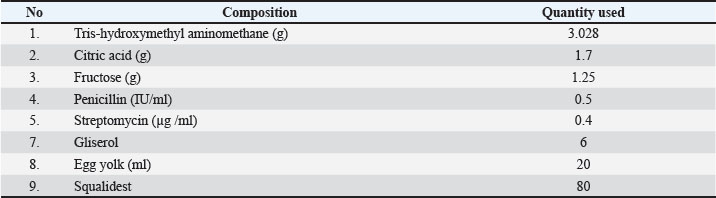
Fig. 1. Workflow of the research procedure, which includes semen collection, fresh semen evaluation, X and Y sperm separation, dilution, semen freezing, thawed sperm analysis, and molecular verification. Sperm sexing was performed using a 5%–10% BSA column gradient following the protocol described by Yendraliza et al. (2025). After incubation for the designated separation times (30, 45, or 60 minutes), two fractions were collected: the upper fraction (putative X-enriched fraction) and the lower fraction (putative Y-enriched fraction). The term “putative” is used because BSA separation is based on differential migration and does not guarantee chromosomal purity. Therefore, molecular verification was conducted to confirm the presence of Y-chromosome–specific SRY amplification in the lower fraction. Following separation, each fraction was centrifuged in Brackett-Oliphant (BO) medium at 1,800 ×g for 10 minutes at 25°C. The pellet was resuspended in 1,000 µl BO medium and evaluated for motility, abnormality, plasma membrane integrity (IPM), and acrosome integrity before cryopreservation. After sexing, semen was diluted using Tris–egg yolk extender (Table 1), equilibrated at 5°C for 4 hours, vapor-frozen, and stored in liquid nitrogen. Post-thaw sperm quality evaluation was performed 24 hours after cryopreservation. Table 1. Tris-Egg Yolk Extender (100 ml) composition.
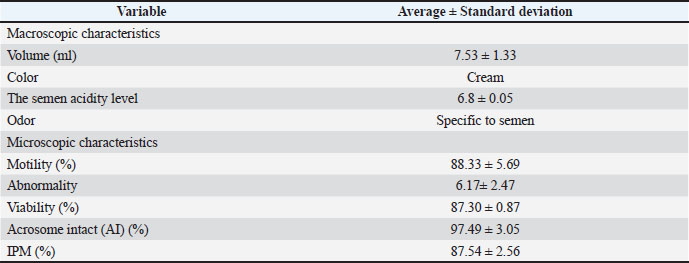
Molecular verificationDNA extractionFor DNA extraction, 0.5 ml of thawed X- and Y-bearing sperm samples were transferred into 1.5 ml microtubes. The samples were centrifuged at 5,000 ×g for 10 minutes at room temperature, after which the supernatant was carefully removed using a 100 µl syringe, leaving approximately 100 µl of sperm pellet in the tube. The pellets were then subjected to DNA extraction using the Geneaid Presto™ Sperm DNA Kit according to the manufacturer’s protocol. The extracted DNA samples were stored at −20°C until further analysis. To verify the presence of DNA, electrophoresis was performed on 1% agarose gel prepared in 100 ml of 1× Tris-Borate-Edta buffer and stained with ethidium bromide. Electrophoresis was performed for 45 minutes at 100 V, 20 W, and 100 mA, and the DNA bands were visualized under UV illumination. X- and Y-sperm amplificationPolymerase chain reaction (PCR) amplification of extracted sperm DNA was performed in a 25 µl reaction mixture containing 2 µl of template DNA, 7.5 µl of master mix, 3 µl of forward and reverse primer mix, and 12.5 µl of nuclease-free water. Two primer sets were used: SRY (SRY-F: 5′-AAGGGGAGAACAGTTAGGGAGAG-3; SRY-R: 5′-ATCGGGTTGCATAGTATTGAAG-3′) and GAPDH (GAPDH-F: 5′-GTGGCGCCAAGAGGGTCATCATC-3′; GAPDH-R: 5′-GGTTTCTCCAGGCGGCAGGT-3′) (Bioneer). Amplification was conducted in a thermocycler with the following profile: initial denaturation at 94°C for 5 minutes; 45 cycles of denaturation at 94°C for 30 seconds, annealing at 60°C for 30 seconds, and extension at 72°C for 30 seconds; followed by a final extension at 72°C for 10 minutes. The PCR products were resolved by electrophoresis on 2% agarose gel stained with ethidium bromide, run at 100 V for 45 minutes, and visualized under UV illumination. Sperm quality assessmentMotility: Sperm motility was evaluated by placing a drop of semen on a glass slide, covering it with a cover slip, and observing it under a light microscope (Motic BA310®) at 450× magnification. The percentage of motile sperm was calculated by multiplying the number of progressively motile spermatozoa by the total spermatozoa observed by 100. Morphological abnormalities: Sperm abnormalities were assessed using eosin smear preparations. In brief, 1 ml of eosin stain was mixed with 0.2 ml of semen, smeared onto a glass slide, and air-dried. The slides were examined under a light microscope (Motic BA310®) at 450× magnification. The evaluation classified spermatozoa as abnormal when they showed curled tails, broken tails, or bent midpieces. The percentage of abnormal sperm was calculated as the number of abnormal spermatozoa divided by the total number of spermatozoa observed, multiplied by 100. IPM: IPM was determined using the hypoosmotic swelling test (HOST). The HOST solution consisted of 1.35 g of fructose and 0.735 g of sodium citrate dissolved in 100 ml of distilled water. A total of 20 µl semen was mixed with 200 µl of HOST solution, gently homogenized, and incubated at 37°C for 45 minutes. After incubation, 10 µl of the mixture was placed on a glass slide, covered with a cover slip, and examined under a light microscope (Motic BA310®) at 400× magnification. Spermatozoa with swollen or coiled tails were classified as having intact plasma membranes, whereas spermatozoa with straight tails were considered to have damaged membranes. The percentage of spermatozoa with intact membranes was calculated as the number of spermatozoa with intact membranes divided by the total spermatozoa observed, multiplied by 100. Acrosome integrity: Acrosome integrity status was evaluated by mixing semen with formal saline solution (physiological NaCl supplemented with 1% formalin) in a microtube at a ratio of 1:5 (Yendraliza and Rahman, 2023). After equilibration, the samples were examined under a phase-contrast microscope (400× magnification), with at least 200 spermatozoa assessed per sample. The presence of a distinct dark-stained region on the sperm head identified spermatozoa with intact acrosomes. Statistical analysisAll statistical analyses were performed using the ejaculate as the experimental unit. Data are presented as mean ± standard error of the mean. The experiment followed a 3 × 3 factorial arrangement in a RCBD, with factor A being the separation time (30, 45, and 60 minutes) and factor B being the bulls’ age (8, 9, and 10 years). Weeks of semen collection were divided into groups. If a significant effect was detected (p < 0.05), the means were separated using Duncan’s multiple range test. The exact p-value is reported in the Results section. Interaction effects are only presented if statistically significant. For molecular verification data (SRY–GAPDH amplification), the results were analyzed descriptively. All statistical analyses were performed using the Statistical Package for the Social Sciences version 25 (IBM Corp. USA). Ethical approvalThis research was approved by the Research Ethics Commission of the Research and Community Service Institute of Universitas Islam Negeri Sultan Syarif Kasim Riau, Riau, Indonesia, in Number 572/Un.04/L.1/TL.01/07/2024. ResultsFresh sperm quality of Bali bullFresh semen quality from Bali cattle. Table 2 shows the average quality of fresh Bali cattle semen after collection. Table 2. Macroscopic and microscopic characteristics of fresh pesisir bull semen.
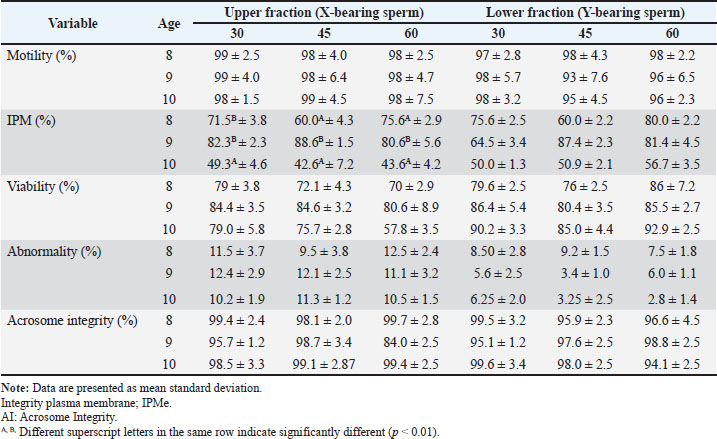
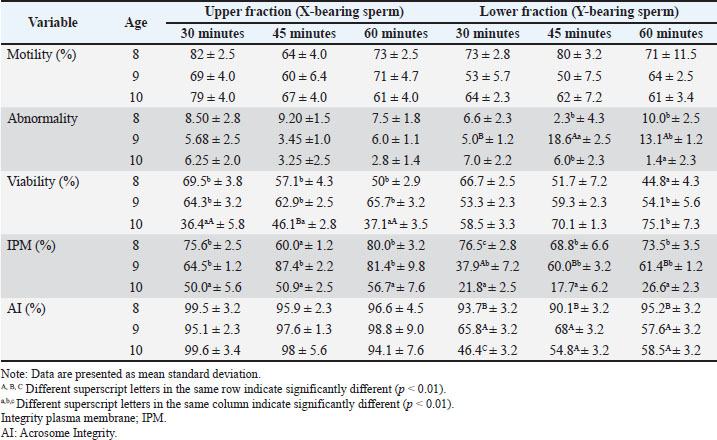
Fresh Bali bull semen exhibited an average volume of 7.53 ± 1.33 ml, a cream color, and a pH of 6.1 ± 0.05. Microscopic analysis showed that sperm motility, viability, IPM, and acrosome integrity were 88.33% ± 5.69%, 87.3% ± 0.87%, 87.54% ± 2.56%, and 97.49% ± 3.05%, respectively. Sperm quality of semen separated before freezingTable 3 presents the average semen quality of Bali cattle sperm after sex separation before freezing. Two-way ANOVA revealed no significant interaction between separation time and bull age for motility in the upper and lower fractions (p > 0.05). Motility was not significantly affected by the separation time or bull age. The viability of the upper fraction was significantly affected by bull age (p < 0.05), but not by separation time (p > 0.05), and no significant interaction was detected (p > 0.05). The mean viability values of 9-year-old bulls were higher than those of 10-year-old bulls, while 8-year-old bulls did not differ significantly from the other age groups. In contrast, viability in the lower fraction was not significantly influenced by separation time (p > 0.05), but the bull age approached significance (p < 0.05). No significant interaction was observed (p > 0.05). The age of the bull significantly affected IPM in the upper fraction (p < 0.01). The highest IPM values were observed in ejaculates from 9-year-old bulls. Separation time did not significantly affect IPM in the upper fraction (p > 0.05), and no significant interaction was detected (p >0.05). IPM in the lower fraction was significantly associated with bull age (p < 0.01), with 9-year-old bulls showing higher IPM than 10-year-old bulls. Separation time had no significant effect on IPM (p > 0.05), and no interaction effect was detected (p > 0.05). Acrosome intact (AI) in the upper fraction was significantly affected by separation time (p < 0.05), but not by bull age (p > 0.05). The 30-minute separation time tended to maintain higher acrosome integrity than the 60-minute separation time. No significant interaction effect was observed (p > 0.05). Acrosome integrity in the lower fraction was significantly influenced by the separation time (p < 0.05), with the 30-minute treatment yielding numerically higher AI values than the longer incubation periods. Bull’s age had no significant effect (p > 0.05), and the interaction was not significant (p > 0.05). A significant interaction between separation time and bull age was detected for the abnormality rate in the upper fraction (p=0.009). Abnormality varied across separation times within specific age groups, indicating that the effect of incubation duration depended on bull age. The abnormality rate in the lower fraction showed a significant interaction effect (p < 0.01), indicating that the response of Y-bearing sperm abnormality to separation time differed among age groups. Table 3. Effect of bull age and separation time on semen quality parameters in the upper and lower fractions of BSA column separation before freezing.
Sperm quality of semen separated after thawingTable 4 presents the average quality of Bali cattle semen from sexing after freezing. The post-thaw motility in the upper fraction was significantly affected by separation time (p < 0.05). The 30-minute treatment maintained higher motility than the 45-minute treatment, while the 60-minute treatment showed intermediate values. Bull’s age did not significantly affect post-thaw motility (p > 0.05), and no interaction effect was observed. Post-thaw motility in the lower fraction was significantly influenced by separation time (p < 0.05), with the 30-minute incubation maintaining higher motility than the 45-minute incubation. The bull’s age had a marginal effect (p=0.067), and no significant interaction was observed. Bull age and separation time significantly influenced the viability of the upper fraction (p < 0.05). Nine-year-old bulls exhibited higher viability than 10-year-old bulls. No significant interaction was detected. The age of the bull significantly influenced the viability of the lower fraction (p < 0.01). Ten-year-old bulls exhibited relatively stable viability values compared with younger bulls. The separation time had a modest but significant effect (p=0.031). No interaction was detected. The IPM in the upper fraction showed a strong main effect of bull age (p < 0.001) and a significant interaction with separation time (p=0.014). The highest IPM values were generally observed in 9-year-old bulls at 45 and 60 minutes, whereas 10-year-old bulls consistently showed lower IPM across separation times. IPM in the lower fraction was strongly influenced by bull age (p < 0.01) and significantly correlated with separation time. Ten-year-old bulls exhibited markedly lower IPM in the lower fraction, particularly at shorter incubation times. Acrosome integrity in the upper and lower fractions was significantly affected by bull age (p < 0.01) but not by separation time. No significant interaction effect was detected. The abnormality rate in the upper fraction did not differ significantly across separation times or bull ages, and no significant interaction was observed (p > 0.05). The abnormality rate in the lower fraction showed a significant interaction effect (p < 0.01), indicating age-dependent responses to separation time. Table 4. Effect of bull age and separation time on semen quality parameters in the upper and lower fractions of BSA column separation after thawing.
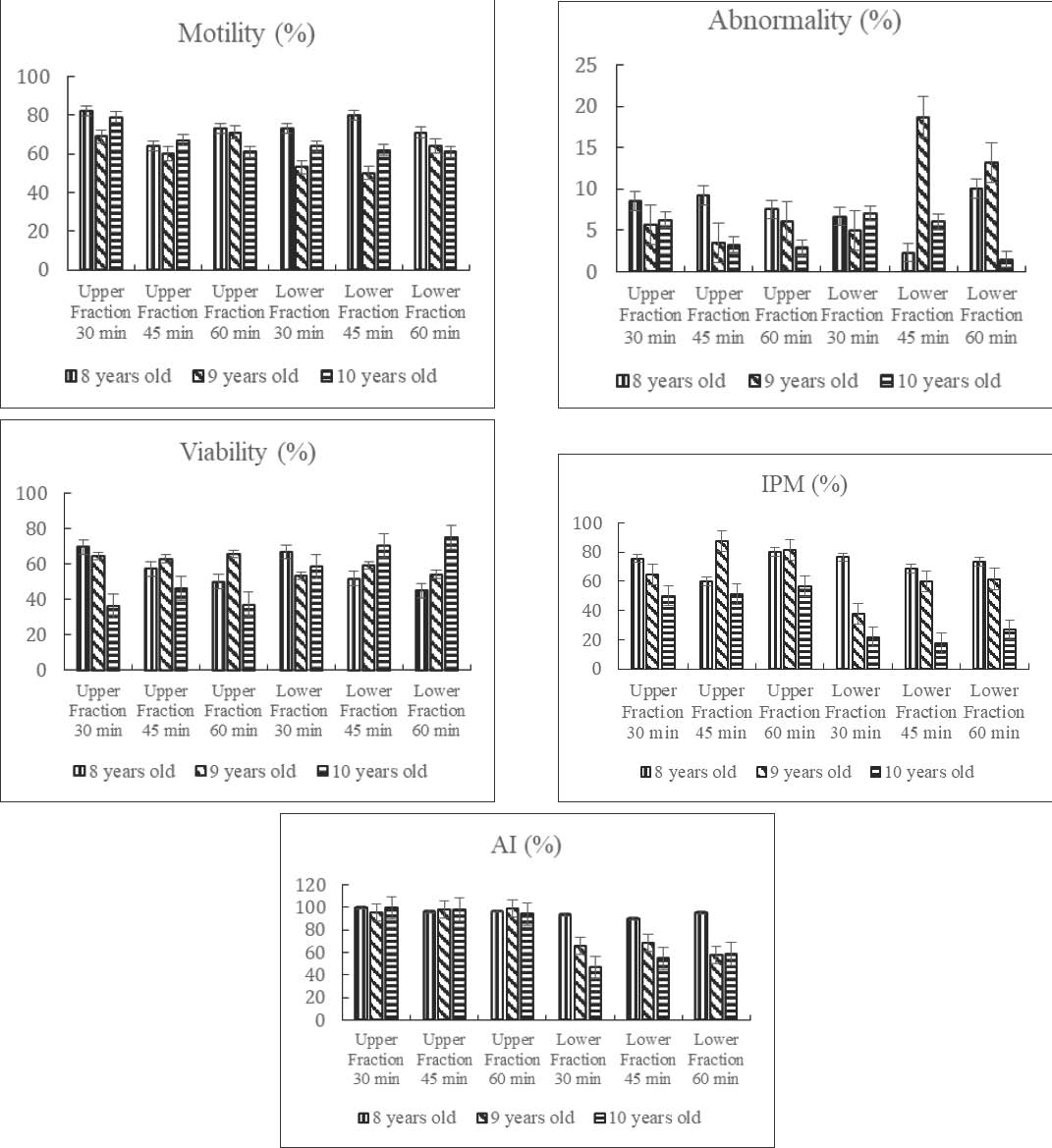
Figure 2 shows the influence of bull age on post-thaw semen quality parameters following BSA-based sperm separation. Semen from 9-year-old bulls consistently exhibited superior motility, viability, and membrane integrity, directly addressing the study objective of determining the most suitable bull age for producing high-quality sexed semen.
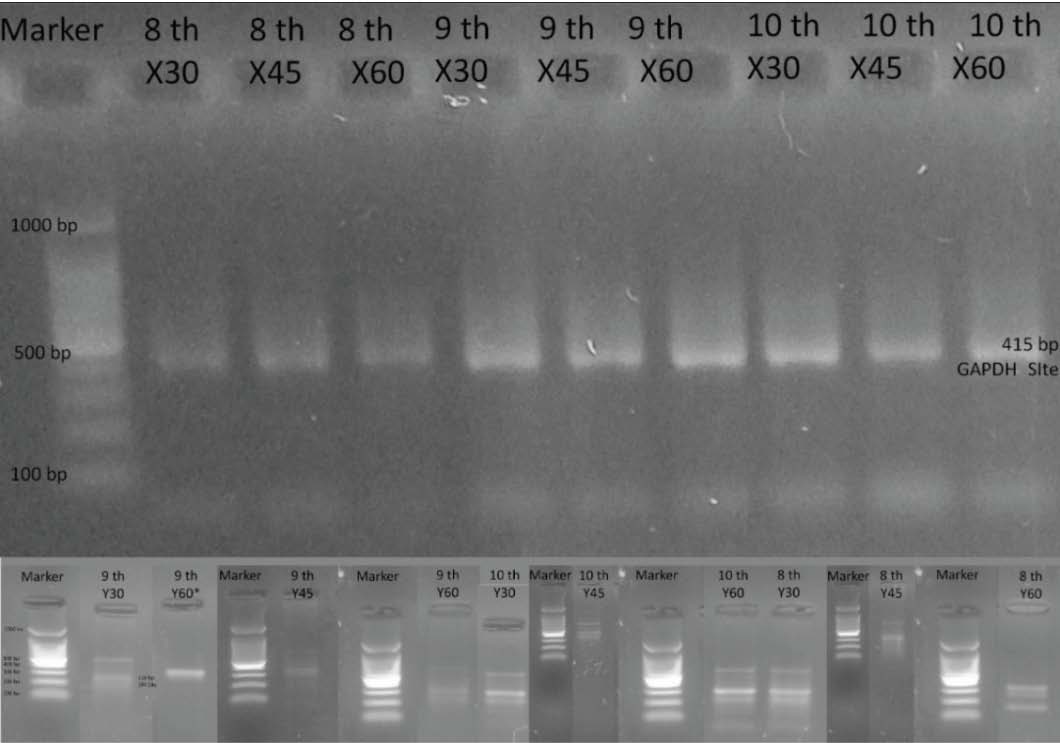
Fig. 2. Effect of bull age and separation time on semen quality parameters in the upper and lower fractions of BSA column separation after thawing. Nevertheless, spermatozoa from the lower fraction of mature bulls maintained relatively stable functional attributes even after extended incubation, highlighting greater resilience compared with that of younger bulls. Verification of sperm sexing with DNA extractionThe molecular verification performed in this study was qualitative, where DNA was detected with clearly visible DNA using the Geneaid Presto™ Sperm DNA Kit. Conventional PCR confirms the presence or absence of Y-chromosome–specific SRY amplification but does not quantify the proportion of X- and Y-bearing spermatozoa. Across separation durations of 30, 45, and 60 minutes and among Bali bulls of different ages, molecular analysis revealed that the upper fraction consistently exhibited a single 415-bp band corresponding to GAPDH, whereas the lower fraction enriched in Y-bearing spermatozoa—displayed two distinct bands at 415 and 318 bp, the latter representing the Y-chromosome–specific SRY marker (Fig. 3).
Fig. 3. Results of amplification of X and Y sperm from the top and bottom layers of Bali cattle semen. Fig. 3 illustrates the effect of separation duration on the molecular discrimination of X- and Y-bearing sperm fractions, as verified by SRY and GAPDH amplification. The results demonstrate that a 30-minute separation provides the clearest differentiation between X and Y fractions, supporting the study objective of identifying the optimal separation time for BSA-based sperm. DiscussionIn this study, macroscopic and microscopic evaluations confirmed that the initial quality of Bali bull semen is high. The average values of motility, abnormality, viability, intact plasma membrane, and acrosome integrity of Bali cattle spermatozoa were eligible to be used as frozen semen under Indonesian national standards for frozen semen (Standar Nasional Indonesia, 2017). The fresh semen rate of Bali bulls was above that of Tuah Sakato bulls (Yendraliza et al., 2020). This was due to the fresh semen from superior Bali bulls, and the good maintenance and feed system given to Balinese cows. Indicate its suitability for AI programs. The current findings reinforce the role of the BSA gradient in enriching sperm populations with superior motility, a mechanism that relies on spermatozoa’s differential migration ability within albumin layers (Vishwanath and Moreno, 2018). In this study, separation time (30–60 minutes) did not significantly affect motility, which aligns with earlier work indicating that BSA separation primarily selects for forward-progressive sperm, and additional incubation beyond optimal thresholds does not further improve separation efficiency (Rasad et al., 2020). Interestingly, the pre-freezing motility values in both X- and Y-bearing fractions of Bali cattle exceeded those reported in buffalo (Asma-ul-Husna et al., 2017) and Sumba cattle (Maulana et al., 2019), suggesting that the efficiency of BSA-mediated selection may be influenced by species-specific seminal plasma composition and sperm membrane architecture. The superior performance of spermatozoa from younger to middle-aged bulls in this study may reflect a more robust antioxidant defense system and greater tolerance to osmotic stress during cryopreservation, consistent with the role of mitochondria in counteracting reactive oxygen species damage (Zoca et al., 2023). This observation is in line with reports that advancing paternal age is associated with increased oxidative stress, reduced mitochondrial activity, and compromised DNA integrity (Aitken et al., 2012; D’Occhio et al., 2020). Intact acrosomes are essential for zona pellucida penetration and oocyte fusion (Aitken et al., 2012), and compromised acrosomes may partially explain reduced fertility outcomes in aged sires. The decline in acrosome stability with age may be linked to cumulative oxidative insults and membrane lipid peroxidation, phenomena well-documented in mammalian sperm physiology (Akhtar et al., 2022). Bali cattle demonstrated relatively higher post-thaw sperm resilience, both in motility and IPM, compared with other livestock species. This supports the hypothesis that breed-specific genetic factors and seminal plasma protein composition contribute to cryotolerance (Sarsaifi et al., 2015). However, confounding factors include variations in methodologies across studies, such as differences in diluents, centrifugation force, and freezing protocols. The swim-up technique, while widely used, often yields lower motility and viability than BSA gradient separation, highlighting the importance of method-specific optimization (Priyanto et al., 2022). Overall, these findings suggest that while the BSA gradient reliably selects high-motility sperm, the ultimate success of sexed semen depends on intrinsic sperm characteristics shaped by bull age and the resilience of sperm membranes to cryogenic stress. DNA was successfully extracted from both upper (X-bearing) and lower (Y-bearing) fractions across all separation times and bull age groups using the Geneaid Presto™ Sperm DNA Kit, with clear and distinct bands observed. These findings are consistent with previous reports on coastal cattle (Mundana et al., 2023), albeit under different processing conditions, and with earlier studies on Simmental cattle, where alternative extraction kits were employed (Kaiin et al., 2017). The results demonstrate that sperm DNA integrity can be reliably preserved across breeds and extraction protocols, reinforcing the robustness of sexed semen as a genomic resource. Among the evaluated incubation periods, 30 minutes proved to be optimal for generating clear Y-sperm DNA amplification, whereas X-sperm DNA could be consistently detected across all separation times. These findings corroborate previous evidence that incubation duration can significantly influence DNA detection efficiency (Nisfimawardah et al., 2023). Notably, phenotypic changes during sperm migration through separation media, as suggested by Taylor et al. (2018), may underlie these differences in amplification clarity. The differential detectability of Y-sperm DNA at shorter incubation intervals highlights the role of BSA gradients in the selective enrichment of sperm subpopulations based on motility and membrane functionality. Similar mechanisms have been observed in density gradient centrifugation, where incubation dynamics modulate sperm recovery efficiency (Vishwanath and Moreno, 2018). The superior amplification at 30 minutes suggests that prolonged incubation may induce structural or oxidative stress that compromises DNA accessibility, a phenomenon previously documented in bovine sperm subjected to extended handling (Kumar et al., 2016). Furthermore, the observed age effect aligns with reports that middle-aged bulls exhibit optimal sperm chromatin stability compared with younger or older sires, likely due to balanced antioxidant defenses and mitochondrial performance (Younus et al., 2024). Taken together, these findings emphasize that both technical parameters (i.e., separation time) and biological factors (i.e., sire age) critically influence the success of sperm and downstream DNA amplification. Optimizing these factors may enhance not only diagnostic accuracy but also the practical application of sexed semen in breeding programs, offering a more sustainable pathway for genetic improvement in Bali cattle and other indigenous breeds. The best separation time for producing clear Y sperm DNA was 30 minutes, while X sperm could be detected with DNA at all separation times. The age of the cattle did not affect the detection of DNA in X and Y sperm, which is consistent with the statement by Nisfimawardah et al. (2023)that separation time can affect DNA detection. Taylor et al. (2018)revealed that the sperm separation process alters the sperm phenotype. X sperm separation time of 30 minutes can detect X sperm and Y sperm DNA more clearly in Balinese cattle semen using BSA column into egg yolk tris diluent which can maintain motility values of 60–82 (X sperm) and 50%–80% (Y sperm), viability of 36%–69% (X sperm) and 53%–75% (Y sperm), intact plasma membrane (50%–87%) X sperm and 17%–76% Y sperm), and acrosome integrity 94%–99% for X sperm and 46%–95% for Y spermatozoa. Age of 9 years produces the best spermatozoa quality. ConclusionThis study demonstrated that separation time during BSA column separation and bull age significantly influenced the semen quality of Bali cattle after thawing. Shorter separation times, specifically 30 minues, consistently produced superior results in terms of DNA detection, motility, viability, membrane integrity, and acrosome preservation. A 9-year-old bull produced the highest-quality post-thaw spermatozoa. These findings provide important insights for refining reproductive biotechnology in Bali cattle, contributing to improved fertility management and the conservation of this valuable native breed. Further studies combining conception rates and sex ratios of offspring are needed to confirm the practical application of these results in field conditions. AcknowledgmentsWe would like to thank Dewi Sartika, Ibna Hayati, and Mr. Kahirul for their contribution. Conflict of interestThe authors declare no conflict of interest. FundingThe authors sincerely thank the Director of Islamic Higher Education, who has funded this research with Decree No. SK 4263 of 2024 for International Collaborative Development Research in 2024. Authors' contributionsYendraliza contributed to the conceptualization and supervision of the study. Muhammad Rodiallah was responsible for the development of the methodology, data collection, and laboratory analysis. Jaswandi performed data analysis, manuscript writing, and interpretation of the results. All authors reviewed and edited the manuscript and approved the final version for submission. Data availabilityAll data are available in the revised manuscript. ReferencesAgus, A. and Mastuti Widi, T.S. 2018. Current situation and future prospects for beef cattle production in Indonesia - a review. Asian-Aust. J. Anim. Sci. 31(7), 976–983; doi:10.5713/ajas.18.0233 Aitken, R.J., Jones, K.T. and Robertson, S.A. 2012. Reactive oxygen species and sperm function-in sickness and in health. J. Androl. 33(6), 1096–1106; doi:10.2164/jandrol.112.016535 Akhtar, M.F., Ma, Q., Li, Y., Chai, W., Zhang, Z., Li, L. and Wang, C. 2022. Effect of sperm cryopreservation in farm animals. Animal 12, 2277. Asma-Ul-Husna, M.A., Mehmood, A., Sultana, T., Shahzad, Q., Ansari, M.S., Rakha, B.A., Saqlan Naqvi, S.M. and Akhter, S. 2017. Sperm sexing in Nili-Ravi buffalo through modified swim up: validation using SYBR®green real-time PCR. Anim. Reprod. Sci. 182, 69–76; doi: 10.1016/j.anireprosci.2017.04.011 BPS. 2024. Populasi Ternak Menurut Kabupaten/Kota Provinsi Riau. Pekanbaru, Indonesia: Badan Pusat Statistik. Available via https://jateng.bps.go.id/statictable/2016/08/23/1317/populasi-ternak-menurut-kabupaten-kota-dan-jenis-ternak-di-provinsi-jawa-tengah-2015.html Butler, S.T., Hutchinson, I.A., Cromie, A.R. and Shalloo, L. 2014. Applications and cost benefits of sexed semen in pasture-based dairy production systems. Anim. Sci. 8, 165–172; doi:10.1017/S1751731114000664 D’Occhio, M.J., Ghuman, S.S., Neglia, G., Della Valle, G., Baruselli, P.S., Zicarelli, L., Visintin, J.A., Sarkar, M. and Campanile, G. 2020. Exogenous and endogenous factors in seasonality of reproduction in buffalo: a review. Theriogenology 150, 186–192; doi:10.1016/j.theriogenology.2020.01.044 Garner, D.L. and Seidel, G.E. 2008. History of commercializing sexed semen for cattle. Theriogenology 69(7), 886–895; doi:10.1016/j.theriogenology.2008.01.006 Kaiin, E.M., Gunawan, M., Octaviana, S. and Nuswantara, S. 2017. Molecular verification of sperm sexing method with BSA (bovine serum albumin) column. Pros. Sem. Nas. Masy. Biodiv. Indonesia 3, 241–245; doi:10.13057/psnmbi/m030214 Kumar, D., Kumar, P., Singh, P., Yadav, S.P. and Yadav, P.S. 2016. Assessment of sperm damages during different stages of cryopreservation in water buffalo by fluorescent probes. Cytotechnology 68(3), 451–458; doi:10.1007/s10616-014-9798-9 Maulana, T., Said, S., Arifiantini, R.I. and Setiadi, M.A. 2019. Sex sorting sperm of sumba Ongole bulls by using snakehead fish (Channa striata) albumin extract. J. Indonesian Trop. Anim. Agric. 44(1), 106–113; doi:10.14710/jitaa.41.1.106-113 Mundana, M., Afriani, T., Yurnalis, Y., Rastosari, A., Oktavianti, F., Al Razak, M.C., Asyraf, M. and Merdana, I.M. 2023. Molecular verification of spermatozoa sexing method in pesisir cattle bull using bovine serum albumin column. Int. J. Vet. Sci. 13, 7–12. Nisfimawardah, L., Firmawati, A., Nur Ihsan, M., Susilawati, T. and Wahjuningsih, S. 2023. Semen cryopreservation quality and sperm kinematics of Saanen goats using different diluents. World’s Vet. J. 13(2), 300–309; doi:10.54203/scil.2023.wvj32 Oldenhof, H., Gojowsky, M., Wang, S., Henke, S., Yu, C., Rohn, K., Wolkers, W.F. and Sieme, H. 2013. Osmotic stress and membrane phase changes during freezing of stallion sperm: mode of action of cryoprotective agents. Biol. Reprod. 88(3), 1–11; doi:10.1095/biolreprod.112.104661 Priyanto, L., Riswandi, R., Putranti, O.D., Gunawan, M., Susanda, A. and Khotimah, K. 2022. The effect of centrifugation time on sexing spermatozoa with bovine serum albumin media on the morphology of X-Y spermatozoa in Simmental cattle. Chalaza J. Anim. Husbandry 7(1), 6–10; doi:10.14334/jitv/v25i3.2494 Purwantara, B., Noor, R., Andersson, G. and Rodriguez‐Martinez, H. 2012. Banteng and Bali cattle in Indonesia: status and forecasts. Reprod. Domestic Anim. 47(Suppl. 1), 2–6; doi:10.1111/j.1439-0531.2011.01956.x Rasad, S.D., Solihati, N., Winangun, K., Yusrina, A. and Avicenna, F. 2020. Effect of incubation time during sperm sexing process on sperm quality of Pasundan bull. Jurnal. Ilmu. Ternak. Dan. Veteriner. 25(3), 112–119; doi:10.14334/jitv/v25i3.2494 Sarsaifi, K., Vejayan, J., Wahid Haron, A., Yusoff, R., Hani, H., Rasoli, M., Ariff Omar, M. and Mazni Othman, A. 2015. Protein profile and functionality of spermatozoa from two semen collection methods in Bali bulls. Livestock Sci. 172, 96–105; doi:10.1016/j.livsci.2014.12.004 Standar Nasional Indonesia, S. 4869-1. 2017. Semen beku - Bagian 1: Sapi bovine (dalam bahasa Indonesia). In Badan Standar Nasional Indonesia, Indonesia, Jakarta. Indonesia: Badan Standardisasi Nasional, pp: 1–4. Taylor, J.F., Schnabel, R.D. and Sutovsky, P. 2018. Identification of genomic variants causing sperm abnormalities and reduced male fertility. Anim. Reprod. Sci. 194, 57–62; doi:10.1016/j.anireprosci.2018.02.007 Vishwanath, R. and Moreno, J.F. 2018. Review: semen sexing-curent state of the art with emphasis on bovine species. Animals 12(1), s85–s96; doi:10.1016/j.animal.2018.09.013 Warman, A.D., Panjono, P., Sawitri, W., Wijanarko, A.W., Atmoko, B.A., Bintara, S., Widi, T.S.M., Jannah, Z.N. and Baliarti, E. 2025. Effect of age and season on fresh semen quality of Bali bulls in Indonesia. Agric. Nat. Resour. 59, 590207. Yendraliza, Y. and Rahman, Y.G. 2023. Cryopreservation of Simmental cattle semen with egg yolk from different avian species and level glycerol of different in tris diluent. Livestock Anim. Res. 21(10), 147–152; doi:10.1016/j.liveracr.2024.04.016 Yendraliza, Y., Handoko, J. and Istiannah, F. 2025. Motility, abnormality and intact plasma membrane of sexed Bali bull sperm in different equilibration time. Buletin Peternakan 49(1), 307–312; doi:10.21059/buletinpeternak.v Yendraliza , Y., Harahap, E.A., Handoko, J., Rodiallah, M. and Arman, C. 2020. Quality of Bali bull cryopreserved sperm using different extenders and equilibration times on pregnancy rate of Bali cows. Songklanakarin J. Sci. Technol. 42(3), 652–659; doi:10.14456/sjst-psu.2020.83 Younus, A.M.D., Yamanaka, T. and Shimada, M. 2024. The protective effects of antioxidants against endogenous and exogenous oxidative stress on bull sperm. In vitro Cell. Dev. Biol. Anim. 60(9), 969–982; doi:10.1007/s11626-024-00944-w Zoca, S.M., Geary, T.W., Zezeski, A.L., Kerns, K.C., Dalton, J.C., Harstine, B.R., Utt, M.D., Cushman, R.A., Walker, J.A. and Perry, G.A. 2023. Bull field fertility differences can be estimated with in vitro sperm capacitation and flow cytometry. Front. Anim. Sci. 4, 1–16; doi:10.3389/fanim.2023.1180975 | ||
| How to Cite this Article |
| Pubmed Style Yendraliza Y, Rodiallah M, Jaswandi J. Optimizing sperm sexing in Bali cattle: Effects of separation time and bull age on BSA gradient efficiency. Open Vet. J.. 2026; 16(5): 2735-2745. doi:10.5455/OVJ.2026.v16.i5.16 Web Style Yendraliza Y, Rodiallah M, Jaswandi J. Optimizing sperm sexing in Bali cattle: Effects of separation time and bull age on BSA gradient efficiency. https://www.openveterinaryjournal.com/?mno=300726 [Access: June 26, 2026]. doi:10.5455/OVJ.2026.v16.i5.16 AMA (American Medical Association) Style Yendraliza Y, Rodiallah M, Jaswandi J. Optimizing sperm sexing in Bali cattle: Effects of separation time and bull age on BSA gradient efficiency. Open Vet. J.. 2026; 16(5): 2735-2745. doi:10.5455/OVJ.2026.v16.i5.16 Vancouver/ICMJE Style Yendraliza Y, Rodiallah M, Jaswandi J. Optimizing sperm sexing in Bali cattle: Effects of separation time and bull age on BSA gradient efficiency. Open Vet. J.. (2026), [cited June 26, 2026]; 16(5): 2735-2745. doi:10.5455/OVJ.2026.v16.i5.16 Harvard Style Yendraliza, Y., Rodiallah, . M. & Jaswandi, . J. (2026) Optimizing sperm sexing in Bali cattle: Effects of separation time and bull age on BSA gradient efficiency. Open Vet. J., 16 (5), 2735-2745. doi:10.5455/OVJ.2026.v16.i5.16 Turabian Style Yendraliza, Yendraliza, Muhamad Rodiallah, and Jaswandi Jaswandi. 2026. Optimizing sperm sexing in Bali cattle: Effects of separation time and bull age on BSA gradient efficiency. Open Veterinary Journal, 16 (5), 2735-2745. doi:10.5455/OVJ.2026.v16.i5.16 Chicago Style Yendraliza, Yendraliza, Muhamad Rodiallah, and Jaswandi Jaswandi. "Optimizing sperm sexing in Bali cattle: Effects of separation time and bull age on BSA gradient efficiency." Open Veterinary Journal 16 (2026), 2735-2745. doi:10.5455/OVJ.2026.v16.i5.16 MLA (The Modern Language Association) Style Yendraliza, Yendraliza, Muhamad Rodiallah, and Jaswandi Jaswandi. "Optimizing sperm sexing in Bali cattle: Effects of separation time and bull age on BSA gradient efficiency." Open Veterinary Journal 16.5 (2026), 2735-2745. Print. doi:10.5455/OVJ.2026.v16.i5.16 APA (American Psychological Association) Style Yendraliza, Y., Rodiallah, . M. & Jaswandi, . J. (2026) Optimizing sperm sexing in Bali cattle: Effects of separation time and bull age on BSA gradient efficiency. Open Veterinary Journal, 16 (5), 2735-2745. doi:10.5455/OVJ.2026.v16.i5.16 |

