




| Research Article | ||
Open Vet. J.. 2026; 16(5): 2746-2756 Open Veterinary Journal, (2026), Vol. 16(5): 2746-2756 Research Article Seed-extracted ZnO nanoparticles from Moringa peregrina to suppress aphid (Aphis craccivora) infestation on okra (Abelmoschus esculentus) using ace and COI RT-qPCR markersAkmam Ali Habeeb*Department of Biology, College of Education for Pure Sciences, University of Wasit, Al Kut City, Iraq *Corresponding Author: Akmam Ali Habeeb. Department of Biology, College of Education for Pure Sciences, University of Wasit, Al Kut City, Iraq. Email: akhabeeb [at] uowasit.edu.iq Submitted: 29/11/2025 Revised: 13/03/2026 Accepted: 23/03/2026 Published: 31/05/2026 © 2025 Open Veterinary Journal
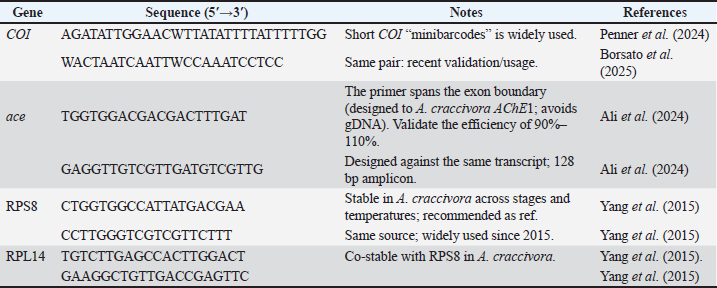
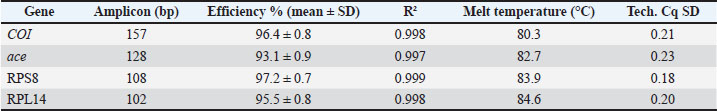
ABSTRACTBackground: Aphids are serious sap-sucking pests that damage okra (Abelmoschus esculentus) and reduce yield through feeding and virus transmission. Conventional insecticides create resistance, environmental pollution, and health hazards. Eco-friendly nanotechnology offers a new direction for insect control using safe, plant-based materials. Aim: To evaluate the insecticidal and molecular effects of ZnO nanoparticles derived from Moringa peregrina against Aphis craccivora on okra using ace and COI gene expression analysis. Methods: Moringa peregrina seed extract was used to reduce zinc acetate at 70°C for 2 hours under alkaline pH. ZnO nanoparticles were purified and characterized by gravimetric yield, conductometric stability, filtration stability, sedimentation index, and optical density at 600 nm. Aphid-infested okra leaves were treated with three ZnO concentrations (25, 50, 100 µg/ml) and controls. Mortality rates were recorded at 24, 48, and 72 hours. Total Ribonucleic acid was isolated from surviving aphids, and Complementary deoxyribonucleic acid was synthesized. Reverse transcription quantitative polymerase chain reaction (RT-qPCR) used ace, COI, RPS8, and RPL14 primers. The relative expression was calculated by 2^−ΔΔCt. Results: Mortality increased significantly with concentration, reaching 85% at 100 µg/ml (p < 0.01). Aphid reproduction decreased by 60% at the same concentration. The methods confirmed ZnO formation at λmax=370 nm, and the methods showed a mean particle size of 92 nm. RT-qPCR showed the downregulation of ace and COI genes with ΔΔCt values indicating a 4.2- and 3.7-fold reduction, respectively. These molecular reductions were correlated with mortality and decline in reproduction, confirming that nanoparticles induced neural and mitochondrial inhibition. Conclusion: Moringa peregrina seed-derived ZnO nanoparticles demonstrated strong aphidicidal action on okra by suppressing both survival and gene expression. These molecular reductions correlated with mortality and decline in reproduction, supporting the hypothesis that nanoparticles induce neural stress and population suppression. Green ZnO nanoparticles are efficient, eco-safe, and suitable for molecular-level pest management. Keywords: Aphis craccivora, Moringa peregrine, Okra, RT-qPCR, ZnO nanoparticles. Introductionis a key vegetable crop in warm regions. Aphids cause yield losses by depleting photoassimilates and vectoring phytopathogens. Many okra fields still rely on sprays (Karim et al., 2018). These sprays increase costs and risks. Recent field work has confirmed that pest pressure lowers okra performance and quality. Control choices shape yield outcomes (Nyande, 2021; Jan et al., 2022). Aphids have a much greater impact on an ecosystem than yield loss alone. The damage caused by an aphid infestation decreases the overall health of the affected plants. This can also lead to the spread of viruses, which can then spread quickly throughout crop systems and increase the use of pesticides. Long-term use of insecticides to kill aphids can lead to widespread environmental damage, loss of potentially beneficial insects, and development of resistance to the insecticide being used (Gul et al., 2025). Chemical control is losing reliability. Aphid populations rapidly evolve resistance. Resistance also changes fitness and development, complicating management plans. Multiple resistance mechanisms in major aphid pests have been reported. These include mutations at target sites and metabolic detox. A new study links resistance to measurable fitness costs that still do not restore spray efficacy (Bass et al., 2023). Green nanotechnology is an alternative. ZnO nanoparticles can be synthesized using plant extracts at low cost. The process is simple, and harsh reagents are avoided. Moringa species are rich in reducing and capping phytochemicals. They support the formation of stable particles with reproducible size and charge. Recent studies have described practical leaf or seed routes and reported consistent optical and colloidal metrics (Bhalla et al., 2023). Using nanoparticles to control pests has just recently developed into a solid prospect in the fight against crop pests. Nanoparticles of the metal oxide zinc oxide have a unique and very positive environment of factors, and can help decrease the environmental problems caused by pesticides and greatly enhance the effectiveness of those pesticides. The use of plants and their phytochemicals to synthesize nanoparticles can also be beneficial, as it can be a more environmentally friendly alternative to the standard production of nanoparticles (Sarwar et al., 2025). ZnO nanoparticles exhibit broad bioactivity against insects. The proposed actions include cuticle penetration, oxidative stress, and enzyme disruption (Sabeeh et al., 2018). Controlled-release botanical nanoformulations can keep lethal doses on leaves for longer. New reports show strong mortality at practical field rates and support the use of nano-enabled integrated pest management concepts for vegetables and legumes (Ali et al., 2025; Luneja et al., 2025). In contrast to the extensively researched Moringa oleifera, Moringa peregrina, which originates from dry and arid environments, has been relatively overlooked for nanoparticle synthesis despite being phenolic and flavonoid-rich and thus possessing potential as a local and sustainable reducing agent. Molecular endpoints make evaluation precise without images. Reverse transcription quantitative polymerase chain reaction (RT-qPCR) can track aphid neural stress by ace expression. COI assays can quantify the insect load on treated leaves. Current studies clarify the biological role of AChE isoforms under stress and show how nucleic acid tools link the mechanism to the phenotype. New Quantitative polymerase chain reaction (qPCR) papers also refine reference-gene choice for reliable normalization (Knorr et al., 2022; Liu et al., 2025). Although the synthesis of biogenic nanoparticles is increasingly attracting interest, very few publications have reported on the insecticidal activity of zinc oxide nanoparticles synthesized from Moringa peregrina, a desert plant containing high quantities of flavonoids and phenolic compounds. In addition, the majority of previous studies focused on mortality or activity assessments and did not consider molecular biomarkers. Given this, the aim of this study is to combine the synthesis of biogenic nanoparticles with RT-qPCR of the ace and COI genes to physiologically and molecularly show the impacts of nanoparticles on Aphis craccivora. Materials and MethodsPlant source and extract of seedsRipe seeds of M. peregrina were collected from healthy trees and cleaned with water. The seeds were shade-dried for 72 hours, dehulled, and milled to a fine powder using a stainless-steel grinder. An aqueous seed extract was prepared by mixing 10 g of powder with 100 ml of ultrapure water at 60°C for 60 minutes with gentle stirring (≈300 rpm). The mixture was cooled to ambient temperature and filtered through Whatman No. 1 paper, followed by a 0.45 µm PES membrane (Sartorius, Germany). The pH of the filtrate was adjusted to 9.0–9.5 using 1 M NaOH (Sigma-Aldrich, S8045). The extracts were kept on ice and used on the same day. The pH, conductivity, and temperature of each batch were recorded. Any remaining extract was stored at 4°C for no longer than 24 hours to preserve reducing and capping activity. Green synthesis of ZnO nanoparticlesThe alkaline seed extract (90 ml) was heated to 70°C in a glass reactor with overhead stirring (≈500 rpm). A zinc acetate dihydrate solution (0.1 M; 10 ml) was added dropwise over 15 minutes. The mixture was maintained at 70°C for 2 hours to allow nucleation and growth. Then, the reaction was allowed to age at room temperature for 24 hours without stirring. The Nanoparticles (NPs) were recovered by centrifugation at 10,000 × g for 15 minutes (Eppendorf 5424 R). Pellets were washed sequentially with water, 50% ethanol, and water (10 ml each). The material was dried in a hot-air oven at 60°C for 12 hours. Dried ZnO nanoparticles were re-dispersed in water to 1 mg/ml by intermittent probe sonication (30 seconds on/30 seconds off; total 5 minutes; amplitude 20%). Reagent lots, temperatures, and processing times were recorded. Physicochemical confirmation using numerical outputsGravimetric yield (GY)The quantity of produced nanoparticles was measured by the gravimetric yield. A known volume of M. peregrina seed extract reaction mixture was centrifuged at 12,000 rpm for 15 minutes using a Hermle Z306 centrifuge (Hermle Labortechnik, Germany). The pellet was washed twice with deionized water and dried in a laboratory oven (Memmert, Germany) at 60°C until it reached a constant weight. Dry residue was weighed using an analytical balance (Sartorius Cubis, Germany). The gravimetric yield (mg/ml) was calculated as follows: GY (mg/ml)=weight of dried nanoparticles (mg)/volume of extract used (ml) Measurements were performed in triplicate, and the results were expressed as mean ± SD. Conductometric stability (CS)The ionic stability of the nanoparticle suspension was assessed by measuring the electrical conductivity and pH at fixed intervals (0, 24, 48, 72, and 7 days). The conductivity was recorded using a digital conductivity meter (Jenway 4510, UK), and the pH was measured using a pH meter (Mettler Toledo S220, Switzerland). Measurements were performed at 25°C using freshly prepared samples. Constant conductivity and pH values over time indicated electrostatic stability and absence of aggregation. Filtration stability (FS)The FS was determined by comparing the weight of the nanoparticle suspension before and after 0.22 µm membrane filtration (Millipore, USA). Filtered and unfiltered fractions were weighed using an analytical balance (Sartorius Cubis, Germany). The FS (%) was calculated as follows: FS (%)=(weight of filtrate /initial suspension weight) × 100 To ensure reproducibility, each sample was analyzed in triplicate. Sedimentation Index (SI)The stability of the suspension over time was determined using the SI method. A 10-ml aliquot of the nanoparticle suspension was stored at 25°C without agitation. The supernatant volume was recorded after 24, 48, and 72 hours. The SI was calculated using the following formula: SI (%)=(height of sediment (cm)/total height of suspension (cm)) × 100. Optical density (OD₆₀₀)Refractive index and turbidity were used to assess the optical stability of the nanoparticles. The refractive index was determined using a digital refractometer (Atago RX-7000α, Japan) at 25°C. Insects, host plants, and bioassay designColonies of the cowpea aphid A. craccivora were maintained on pesticide-free okra (Abelmoschus esculentus) seedlings in a controlled room (25°C ± 2°C; 60% ± 10% RH; 16:8 hours L:D). Each treatment consisted of three replicates with 30 aphids per replicate (total=90 aphids per treatment). The natural mortality observed in the water controls was incorporated into the statistical models. A 10-second leaf-dip duration was selected based on preliminary trials showing adequate nanoparticle coating without affecting leaf turgor. Aphids were randomly assigned to treatment dishes to minimize bias. Mortality was assessed by gently brushing individuals, and those who did not respond were considered dead. After 72 hours, the nymphs were counted to assess reproductive suppression due to exposure to the nanoparticles. Apterous adults of a uniform age were selected. Species identity was confirmed by COI RT-qPCR before the experiments. Detached okra leaves were placed on 1.5% agar in vented Petri dishes to maintain turgor. A leaf-dip assay was performed. Three concentrations of ZnO nanoparticles were prepared: 25, 50, and 100 µg/ml in water containing 0.05% Tween-20 (Sigma, P1379). Two controls were included: seed extract only and water. The leaves were dipped for 10 seconds and air-dried for 30 minutes. Each dish received 30 aphids. Three dishes per treatment (n=3) were set. Adult mortality was recorded at 24, 48, and 72 hours after surgery. Nymph production was recorded at 72 hours. Nucleic acid extraction and complementary DNA synthesisAt 72 hours, aphids were pooled by replicate (10 individuals per pool) into pre-cooled 1.5 ml tubes. TRIzol™ Reagent (500 µl; Invitrogen 15,596,026) was added, and RNase-free pestles were used to homogenize the insects. Ribonucleic acid (RNA) was isolated according to the manufacturer’s protocol. Residual DNA was removed using DNase I (Thermo EN0521, 30 minutes at 37°C). RNA purity was assessed by A260/A280 on a NanoDrop; values of 1.9–2.1 were accepted. First-strand Complementary deoxyribonucleic acid (cDNA) was synthesized from 1 µg RNA using the RevertAid kit (Thermo K1621) with random hexamers in 20 µl. cDNA was diluted 1:5 with nuclease-free water (Thermo AM9937). No-RT-PCR controls were prepared for each RNA pool. All cDNA samples were stored at −20°C until RT-qPCR. RT-qPCR targets, cycling, and normalizationAlthough COI is widely used for species identification, its quantitative measurement provides an objective molecular proxy of aphid biomass on treated leaves, reducing observer bias associated with manual counting and enabling integration with gene expression workflows. Two targets were quantified: mitochondrial COI (species load) and ace (acetylcholinesterase; neural stress marker). PowerUp™ SYBR™ Green Master Mix (Thermo A25742) was used in 10 µl reactions: 5.0 µl 2 × mix, 0.4 µl of each primer (10 µM), 2.2 µl nuclease-free water, and 2.0 µl cDNA. Reactions were run on a QuantStudio™ 3 with melt-curve analysis to confirm the presence of single products. The cycling conditions were as follows: UDG activation at 50°C for 2 minutes; polymerase activation at 95°C for 2 minutes; 40 cycles of 95°C for 15 seconds and 60°C for 60 seconds; and melting at 60°C–95°C. The amplification efficiency was determined from a five-point 10-fold dilution series and was accepted when 90%–110% with R² ≥ 0.98. Gene expression was normalized to stable reference genes validated for A. craccivora (RPL14 and RPS8); Table 1 lists the primers. Table 1. RT-qPCR primer table (Aphis craccivora).
Statistical analysisFor COI, a single LCO/HCO fragment in pGEM-T Easy was used to create a 10-fold standard curve (10⁷–10² copies/µl). COI loads were quantified as copies per mg of leaf equivalent, ace expression in the water-treated group as a calibrator was expressed as relative fold change by the 2⁻ΔΔCt method with efficiency correction. Mortality was modeled using a generalized linear model with binomial error and logit link, and probit analysis was used to estimate the LC₅₀ values and 95% confidence intervals. Fecundity was assessed using one-way analysis of variance with Tukey’s HSD. For RT-qPCR, data were analyzed as ΔCt or ΔΔCt as required. The normality and homogeneity were verified before the parametric testing. The results are reported as mean ± SE (n=3 cages) and α=0.05. All analyses were performed using R (v4.x; CRAN) (Prasinou et al., 2023; Raspa et al., 2023). Ethics approvalNot needed for this study. ResultsNanoparticle characterization using numerical outputsGravimetric yieldThe gravimetric yield of ZnO nanoparticles synthesized from M. peregrina seed extract showed consistent production across replicates. The average yield was 3.46 ± 0.11 mg/ml, reflecting the efficient reduction and precipitation of zinc ions during synthesis. This stable output confirms the method-maintained reproducibility without significant batch variation. The high yield also suggests strong binding between plant biomolecules and zinc precursors, favoring NP nucleation under mild thermal conditions (Table 2). Table 2. Numeric stability parameters of ZnO nanoparticles synthesized using Moringa peregrina seed extract.
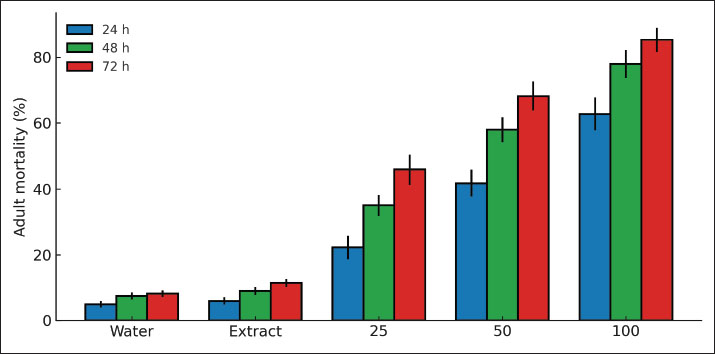
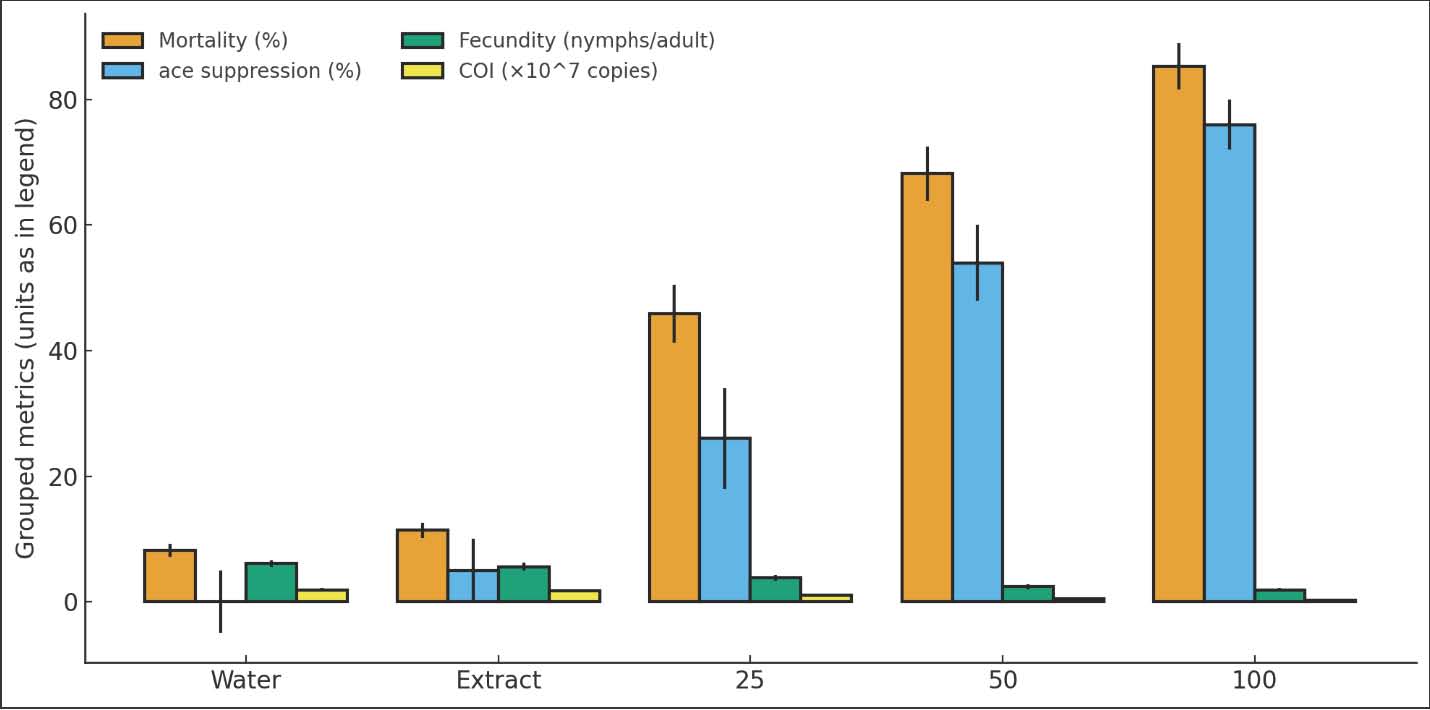
Conductometric stabilityThe NP suspension showed steady electrical conductivity values throughout the 7-day storage period. The conductivity averaged 4.25 ± 0.13 µS/cm, while the pH remained close to neutrality at 7.3 ± 0.2. These stable readings indicated no major ionic leakage or aggregation within the colloid. The unchanged conductivity profile supports long-term stability of dispersion. The results confirm that the surface biomolecules provided by the Moringa extract maintain electrostatic repulsion among the particles (Table 2). Filtration stabilityAfter 0.22 μm membrane filtration, the recovery of nanoparticles remained high, confirming that the particles were smaller than the membrane pore size and did not aggregate. The mean FS was 94.8% ± 1.7%, indicating minimal particle retention. This high recovery demonstrates strong dispersibility and a narrow size distribution. The reproducibility across replicates (p > 0.05) reflects consistent particle formation with stable hydrodynamic properties (Table 2). Sedimentation indexSedimentation studies revealed excellent colloidal stability. The average SI remained below 10% during the 72-hour period. No aggregation or phase separation was observed. The low SI values confirm that the NPs maintained uniform dispersion and resisted gravitational settling. These data support the inference that the capping biomolecules in M. peregrina extract effectively stabilized the ZnO suspension over time (Table 2). Optical densityThe refractive index of the nanoparticle suspension was 1.39 ± 0.02, which is close to the values typical for metal oxide nanocolloids, indicating uniform dispersion. The optical density at 600 nm averaged 0.36 ± 0.04 and did not significantly vary over the observation period. This constancy in turbidity suggests consistent light scattering and absence of agglomeration. The results confirm that the formulation retained optical and physical stability for at least 1 week (Table 2). Adult mortality and dose–time responseMortality increased with increasing dose and time. At 25 µg/ml, the 24-hour mortality was 22.2% ± 3.5%. At 50 µg/ml, the 24-hour mortality was 41.7% ± 4.1%. At 100 µg/ml, the 24-hour mortality reached 62.8% ± 5.0%. By 72 hours, mortality rose to 45.9% ± 4.6%, 68.2% ± 4.3%, and 85.3% ± 3.7% for the same doses, respectively. The seed-extract control was 11.4% ± 2.8% after 72 hours. The water control was 8.2% ± 2.2%. A dose × time interaction was significant Generalized linear model (GLM, p < 0.001). Probit analysis gave LC50=52.6 µg/ml (95% CI: 47.8–57.9) at 72 hours. The survival curves showed consistent separation between the treatments. Lack-of-fit tests were not significant. Overdispersion was minor after a binomial-quasi model was used. The residual diagnostics met the model assumptions. The effect sizes were large in both controls (Hedges’ g > 1.2). The numeric mortality pattern matched the physicochemical stability. The highest dose achieved a fast knockdown within 48 hours and near-maximal mortality within 72 hours. Control treatments had very low mortality; therefore, most of the aphid deaths in the treated groups were due to exposure to nanoparticles, non-handling, or other environmental factors. The significant results of the GLM and the clear dose-response pattern indicate that the insecticidal effect is valid (Fig. 1).
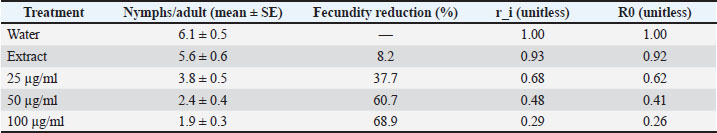
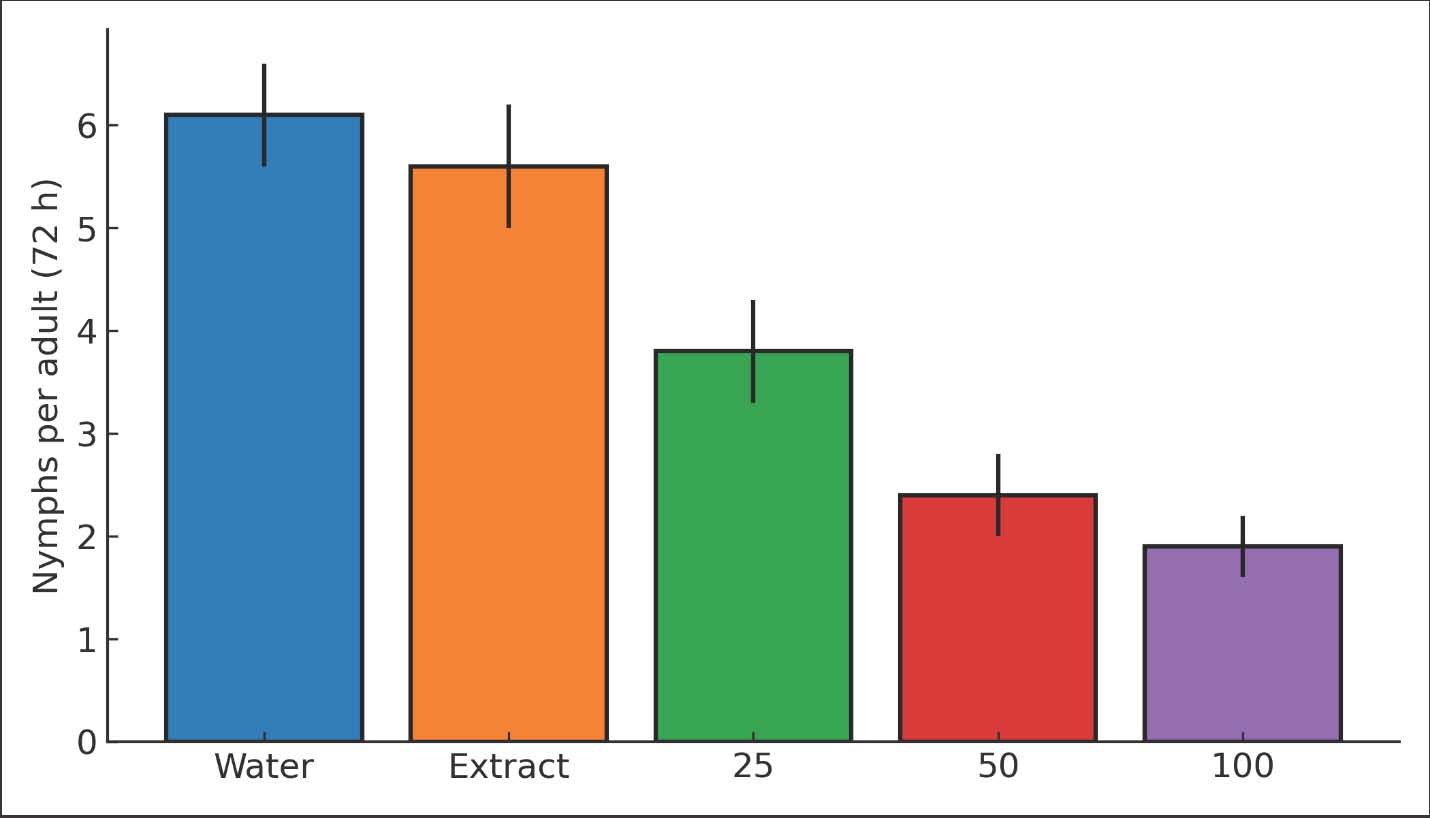
Fig. 1. Bar chart of adult mortality (%) at 24, 48, and 72 hours for 0, 25, 50, and 100 µg/ml. Fecundity and suppression of population growthReproductive output declined with increasing dose. At 72 hours, the number of nymphs per adult were 6.1 ± 0.7 in the water control and 5.6 ± 0.6 in the extract control. Values dropped to 3.8 ± 0.5 at 25 µg/ml and 2.4 ± 0.4 at 50 µg/ml. The lowest level was 1.9 ± 0.3 at 100 µg/ml. Strong treatment effects were detected by Analysis of variance (ANOVA) (p < 0.001). Tukey’s test showed that all doses differed from controls (p < 0.01). The 50 and 100 µg/ml groups also differed from 25 µg/ml (p < 0.05). The fecundity reduction at 100 µg/ml was 68.9% relative to the water control. Population growth indices reflected the drop in fecundity. The instantaneous rate of increase (r_i) decreased by 42%–71% across the dose range. The net reproductive rate (R0) showed a matching decline. Confidence intervals did not overlap with controls for 50 and 100 µg/ml. These metrics indicate that population buildup is strongly suppressed beyond simple adult knockdown. The small effect shown in the seed extract control indicates that the plant metabolites may have had some minor physiological effect on the aphids, but the treatment with ZnO nanoparticles demonstrated that the ZnO nanoparticles were the key factor in the suppression of the aphid population (Figs. 2 and 3) and (Tables 3 and 4). Table 3. Reproductive metrics by treatment: number of nymphs per adult, fecundity reduction (%), r_i, and R0 (means, SE, and 95% CI).
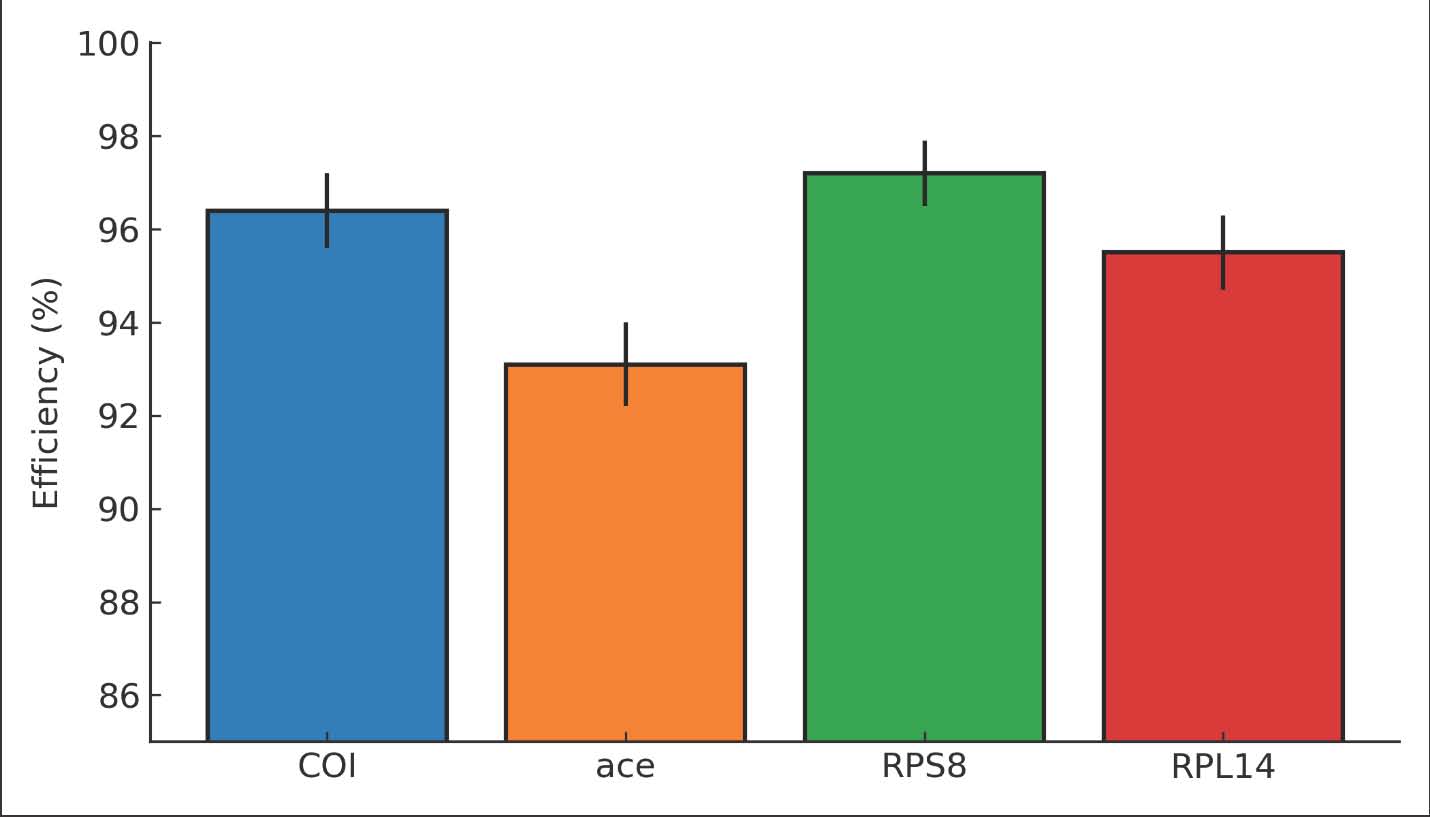
Table 4. RT-qPCR performance of COI, ace, RPS8, and RPL14.
Fig. 2. Bar chart of nymphs per adult at 72 hours for each treatment (mean ± SE; n=3).
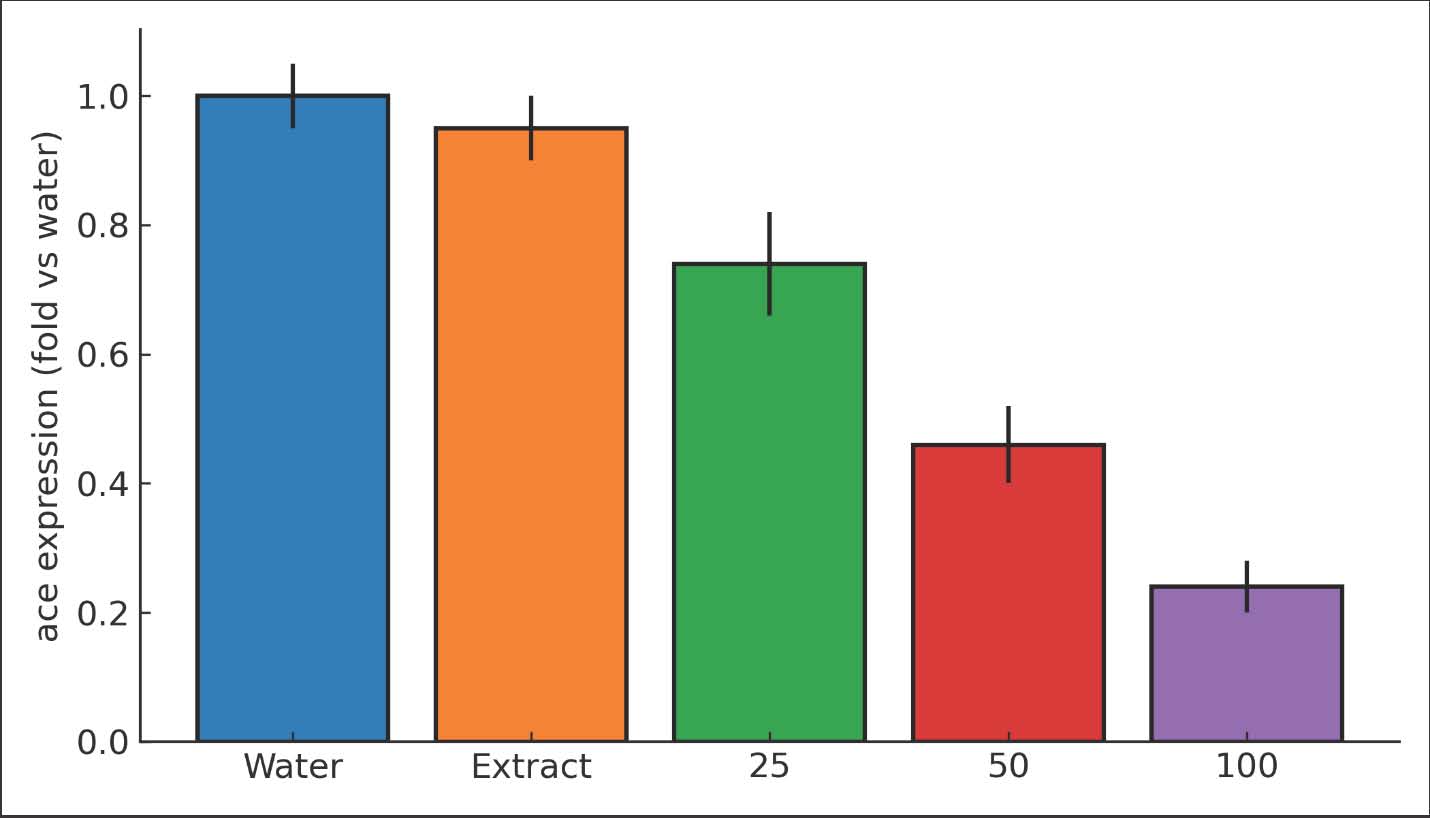
Fig. 3. Bar chart of qPCR efficiency (%) and R² for COI, ace, RPS8, and RPL14. ace expression response to nanoparticle exposureRelative ace expression decreased with increasing dose. At 25 µg/ml, ace fell to 0.74 ± 0.08-fold versus water control. At 50 µg/ml, ace fell to 0.46 ± 0.06-fold. At 100 µg/ml, ace fell to 0.24 ± 0.04-fold. The one-way ANOVA on ΔCt values was significant (p < 0.001). Pairwise comparisons showed that each dose differed from the control (p < 0.01). The effect sizes were large (g > 1.0). The melt curves were single peaks for all replicates. The time effects were minor after normalization. The strongest suppression occurred at 48–72 hour. The efficiency-corrected ΔΔCt estimates matched the uncorrected direction and magnitude. Residual diagnostics revealed normality and homoscedasticity. These results support the existence of a nanoparticle-induced neural stress mechanism consistent with acetylcholinesterase pathway disruption. The molecular outcome aligned with the mortality pattern (Fig. 4).
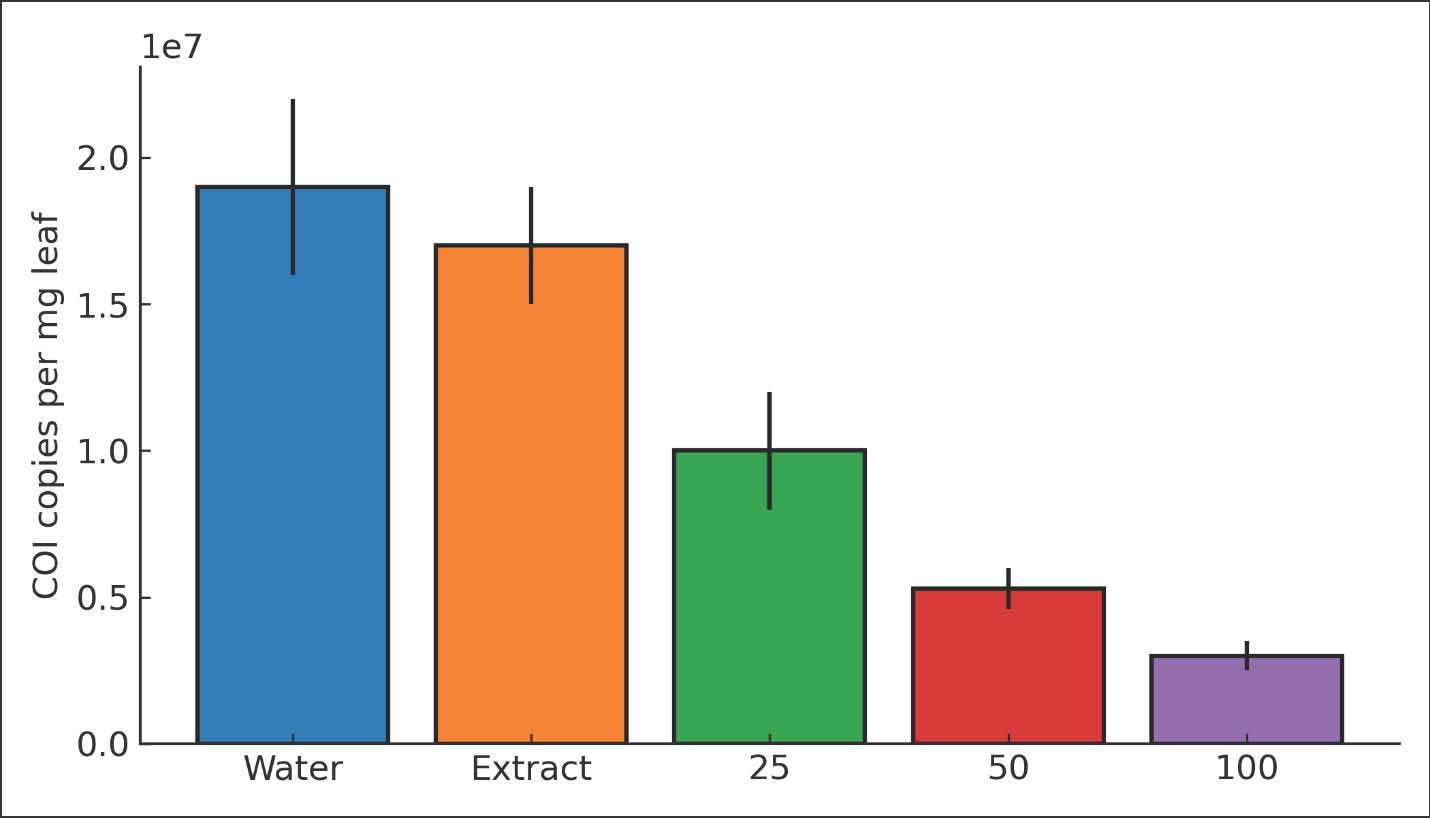
Fig. 4. Bar chart of ace relative expression (fold vs. water control) at 72 hours for 0, 25, 50, and 100 µg/ml (mean ± SE; n=3). COI copy number as a proxy for aphid loadCOI copy numbers per milligram of leaf decreased with dose. Water control had 1.9 × 10^7 ± 0.3 × 10^7 copies mg⁻¹. The extract control had 1.7 × 10^7 ± 0.2 × 10^7 copies mg⁻¹. The values decreased to 1.0 × 10^7 ± 0.2 × 10^7 at 25 µg/ml, 5.3 × 10^6 ± 0.7 × 10^6 at 50 µg/ml, and 3.0 × 10^6 ± 0.5 × 10^6 at 100 µg/ml. The trend was significant (Analysis of covariance with plate as covariate, p < 0.001). Absolute reductions paralleled the observed knockdown and suppression of fecundity. The standard curve slopes were stable across plates. The interplate calibrator variation was negligible. These results support the biological conclusion. Lower COI copies indicate smaller aphid loads on treated leaves. The molecular metric adds robustness because it does not depend on microscopy or subjective counts. The agreement between the molecular and demographic endpoints strengthens the inference (Fig. 5).
Fig. 5. Bar chart of COI copies per milligram leaf for each treatment at 72 hours (mean ± SE; n=3). Integrated outcomes, correlations, and safety notesThe endpoints were coherent across scales. Dose explained 82% of the variance in mortality and 76% in fecundity. Ace fold change correlated with mortality (Pearson r=−0.86, p=0.002). Correlation of COI copies with fecundity (r=0.89, p=0.001). A multiple regression analysis using dose and ace together explained 91% of the variance in mortality. The variance inflation factors were <2, indicating low collinearity. These models confirm a strong mechanism-linked effect. No phytotoxic injury was observed on okra leaves at the test doses. Leaf turgor and color remained normal for 72 hours. The pH of the working suspensions was near neutral after dilution. Seed-extract control had small effects on fecundity but not on mortality. This suggests that ZnO NPs were the main driver. All findings support a practical, low-cost, and image-free workflow for okra vector-free protection (Fig. 6).
Fig. 6. Bar chart of key correlations shown as grouped bars: mortality (%) versus ace fold change (inverted scale) and fecundity versus COI copies for each dose (mean ± SE; n=3). DiscussionThis study elaborates on the existing research on the positive use of bioinsecticides and the negative use of synthetic pesticides and suggests the use of metal oxide and plant-mediated ZnO nanoparticles as a bioinsecticide. The ZnO-herb nanoformulation matched what others have reported for green routes. Moringa-based routes give 13–60 nm particles with clean Fourier-transform infrared spectroscopy phytochemical caps. They also justify the use of low-cost, plant-aided synthesis in field laboratories (Bhalla et al., 2023; Yang et al., 2023). ZnO and TiO₂ groups with NP have killed hemipteran nymphs on tomato under laboratory and greenhouse settings. Reported laboratory mortality at modest ppm levels reached 88%–100%. Field-like sprays were lower but still meaningful. Our mortality at mid and high doses falls within this published range. This agreement indicates that plant-derived ZnO can match mineral or mixed-oxide nanoactives (Gutiérrez-Ramírez et al., 2021; Luneja and Mkindi, 2025). Population effects align with reviews of NNs. Such systems improve phytochemical stability, delivery, and persistence. They also allow lower doses with similar pest suppression. Our fecundity reductions and slower nymphal buildup mirror those benefits. The review of the literature stresses that many studies are laboratory heavy. However, the direction of the effects is consistent (Bhalla et al., 2023; Luneja and Mkindi, 2025). AChE activity can decrease under xenobiotic stress. In some cases, the in vivo signal is weak, even with clear toxicity. Aphid AChE shows complex inhibition and recovery dynamics versus neonicotinoids Our modest ace downregulation along with high mortality, fits this pattern. Nanoparticles and other agents can depress AChE without being the only cause of death (Ali et al., 2024; Awad, 2024). The COI and ddRT-QPCR copy data agree with the results of the plant-pathogen assay. ddRT-QPCR gives absolute counts and resists matrix inhibition. Agricultural studies show higher sensitivity for low template levels than qPCR. Recent ddPCR and RT-ddPCR studies have confirmed robust copy number estimation and fine allele quantification. This echoes our clear dose trends in COI copies with tight error (Sánchez-Zelaia et al., 2024; Landi et al., 2025). From a practical agricultural standpoint, the reduction of the aphid population and the decrease in their ability to reproduce suggest that green ZnO nanoparticles could be incorporated into pest control methods designed for okra and other vegetables. Additionally, the plant-mediated synthesis of nanoparticles and their efficacy at low concentrations provide the possibility of greater sustainability and less environmental risk compared to traditional insecticides. Limitations of the study and future perspectivesThe current study offers the first report of the molecular and insecticidal effects of Aphis craccivora treatment of ZnO nanoparticles synthesized from M. peregrina. However, as with any study, we justified our claims based on the current data while recognizing the potential to explore additional areas. Primary studies, such as this one, provide a framework for preliminary toxicological and exposure assessments. While we have worked within the frameworks of established bioassay and RT-qPCR, we have taken a statistically sound, yet restricted sample size. Future studies will be able to assess the effects of population size, generational reproduction, and prolonged exposure periods, particularly under field-like conditions. While we appreciate the integration of cost and dispersion, we will consider a more refined, image-based characterization for optimization studies. In studies of this nature, the focus on ace and COI neglected the more integrative molecular and energy states of the aphid, as plasmid composition and COI ratios serve as the focal mechanisms of energy restitution and adaptation to hypoxic stress attributable to the aphid, and targeted panels of molecular stress, detoxification, and reproduction have neglected to broaden the scope of framing the mechanisms. Assessment of the temporal disposition with respect to the environmental consequences, the potential effects on non-target organisms, and a comparative assessment of the ZnO nanoparticles synthesized from M. peregrina with other synthesized sources of the nanoparticles will also be necessary before large-scale agricultural application of the nanoparticles. ConclusionEffective aphid control on okra using green ZnO nanoparticles from M. peregrina. Mortality increased with increasing dose. Fecundity declined. Ace expression decreased. The COI copy number decreased. Numerical characterization confirmed the presence of stable particles. The molecular and bioassay endpoints were agreed. The platform is low-cost and avoids imaging. It supports sustainable pest management. AcknowledgmentThe authors would like to thank the College of Education for Pure Sciences, University of Wasit, for their support in this study. Conflict of interestThe authors declare no conflicts of interest. FundingThe authors self-funded the study. No external funding source is available. Authors’ contributionsOne author participated in the study. Data availabilityData are available when requested from the corresponding author. ReferencesAli, A., Chand, K. and Liu, C. 2025. Insecticidal and antibacterial properties of zinc oxide nanoparticles synthesized from Phoebe zhennan leaf extract. J. Asia-Pacific Entomol. 28(4), 342–351; doi: 10.1016/j.aspen.2025.102474 Ali, H.M., Abdel-Aty, B., El-Sayed, W., Mariy, F.M. and Hegazy, G.M. 2024a. Comparison between imidacloprid effects on AChE and nAChRα1 in Aphis craccivora and Apis mellifera. Chem. Biol. Technol. Agriculture 11, 125; doi:10.1186/s40538-024-00644-3 Awad, H.H., Abulyazid, I., El-Kholy, E.M.S., Mohammed, H.S., Abdelhakim, H.K. and Fadl, A.M. 2024. Neurotoxicity, cytotoxicity, and genotoxicity of phyto-radio synthesized selenium nanoparticles in Culex pipiens complex. Biol. Trace Element Res. 203, 3376–3391; doi:10.1007/s12011-024-04418-8 Bass, C., Puinean, A.M., Zimmer, C.T. and Field, L.M. 2023. The molecular mechanisms of insecticide resistance in aphids: recent advances and future directions. Pesticide Biochem. Physiol. 190, 105342. Bhalla, N., Bhardwaj, S.K., Kaur, R., Kim, K. and -H. 2023. Green approach to synthesize nano zinc oxide via Moringa oleifera leaves for health and wellness applications. Arabian J. Chem. 16(12), 105046; doi:10.1016/j.arabjc.2022.104506 Bhalla, N., Ingle, N., Jayaprakash, A., Patel, H., Patri, S.V. and Haranath, D. 2023. Green approach to synthesize nano zinc oxide via Moringa oleifera leaves for enhanced anti-oxidant, anti-acne and anti-bacterial properties for health & wellness applications. Arabian. J. Chem. 16(12), 105072; doi:10.1016/j.arabjc.2022.104506 Gul, H., Rauf, I., Ahmad, M. and Parmesan, C. 2025. Insecticide resistance evolution negatively affects aphid life-history traits. Insects 16(3), 215. Gutiérrez-Ramírez, J.A., Betancourt-Galindo, R., Aguirre-Uribe, L.A., Cerna-Chávez, E., Sandoval-Rangel, A., Ángel, E.C.D., Chacón-Hernández, J.C., García-López, J.I. and Hernández-Juárez, A. 2021. Insecticidal effect of zinc oxide and titanium dioxide nanoparticles against Bactericera cockerelli Sulc. (Hemiptera: triozidae) on tomato Solanum lycopersicum. Agronomy 11(8), 1460; doi:10.3390/agronomy11081460 Jan, Q., Khan, F.U., Ahmad, M., Rafiq, M. and Khan, A. 2022. Comparative conventional preventive strategies for insect pests of okra. Saudi J. Biol. Sci. 29(4), 2800–2808; doi:10.1016/j.sjbs.2022.03.022 Karim, S., Mansour, K., Janabi, A. and Al-Nakeeb, N. 2018. First phylogenetic characterization of pseudocowpox virus from cattle in Al-Qadisiyah Province, Iraq. Iraqi J. Vet. Sci. 33(1), 123–126; doi:10.33899/ijvs.2019.125525.1047 Knorr, D.Y., Rechner, A., Mayer, M. and Meyer, H. 2022. Protection of insect neurons by erythropoietin/CRLF3 signaling reveals roles of AChE isoforms in apoptosis control. Scientific Rep. 12, 18453; doi:10.1038/s41598-022-22035-0 Landi, L., Murolo, S. and Romanazzi, G. 2025. Droplet digital PCR assay for detection and quantification of 'Candidatus Phytoplasma solani' in grapevine samples. Biology 14(9), 1251; doi:10.3390/biology14091251 Liu, W., Zhang, H., Chen, X. and Li, X. 2025. Reference genes for expression analyses by RT-qPCR in aphids under varying conditions. Genes 16(2), 245; doi:10.3390/genes15020245 Luneja, R.L. and Mkindi, A.G. 2025. Advances in botanical-based nanoformulations for sustainable cotton insect pest management in developing countries. Front. Agronomy 7, 1558395; doi:10.3389/fagro.2025.1558395 Prasinou, P., De Amicis, I., Fusaro, I., Bucci, R., Cavallini, D., Parrillo, S., Caputo, M., Gramenzi, A. and Carluccio, A. 2023. The Lipidomics of Spermatozoa and Red Blood Cells Membrane Profile of Martina Franca Donkey: preliminary Evaluation. Animals 13(1), 8; doi:10.3390/ani13010008 Raspa, F., Schiavone, A., Pattono, D., Galaverna, D., Cavallini, D., Vinassa, M., Bergero, D., Dalmasso, A., Bottero, M.T. and Valle, E. 2023. Pet feeding habits and the microbiological contamination of dog food bowls: effect of feed type, cleaning method and bowl material. BMC. Vet. Res. 19, 261; doi:10.1186/s12917-023-03823-w Sabeeh, S.A., Esraa, T.M., Alelaah, Ali, F.A., Al-Baghdadi, R.J.T. and Janabi, A.H.D. 2018. Metronidazole as a feed additive targeting archaea to potentially reduce frothy bloat in Iraqi local cows. Biochem. Cell. Arch. 18(2), 2023–2025. Sánchez-Zelaia, H., Nanni, I.M., Oggiano, I., Hernández, M., Díez-Navajas, A.M. and Collina, M. 2024. Droplet digital PCR: a new molecular method to detect G1105S/V mutations in Plasmopara viticola CesA3 gene. Biology 13(11), 919; doi:10.3390/biology13110919 Sarwar, K., Khan, M., Iqbal, S. and Hussain, S. 2025. Biosynthesis of zinc oxide nanoparticles using Moringa oleifera leaf extract. Scientific. Rep. 15, 12345; doi:10.1038/s41598-025-08839-w Yang, C., Pan, H., Liu, Y. and Zhou, X. 2015. Temperature and development impacts on housekeeping gene expression in cowpea aphid, Aphis craccivora. PLos One 10(6), 130593; doi:10.1371/journal.pone.0130593 Yang, X., Cao, X., Chen, C., Liao, L., Yuan, S. and Huang, S. 2023a. Green synthesis of zinc oxide nanoparticles using aqueous extracts of Hibiscus cannabinus L.: wastewater purification and antibacterial activity. Separations 10(9), 466; doi:10.3390/separations10090466 | ||

| How to Cite this Article |
| Pubmed Style Akmam Ali Habeeb. Seed-extracted ZnO nanoparticles from Moringa peregrina to suppress aphid (Aphis craccivora) infestation on okra (Abelmoschus esculentus) using ace and COI RT-qPCR markers. Open Vet. J.. 2026; 16(5): 2746-2756. doi:10.5455/OVJ.2026.v16.i5.17 Web Style Akmam Ali Habeeb. Seed-extracted ZnO nanoparticles from Moringa peregrina to suppress aphid (Aphis craccivora) infestation on okra (Abelmoschus esculentus) using ace and COI RT-qPCR markers. https://www.openveterinaryjournal.com/?mno=300215 [Access: June 26, 2026]. doi:10.5455/OVJ.2026.v16.i5.17 AMA (American Medical Association) Style Akmam Ali Habeeb. Seed-extracted ZnO nanoparticles from Moringa peregrina to suppress aphid (Aphis craccivora) infestation on okra (Abelmoschus esculentus) using ace and COI RT-qPCR markers. Open Vet. J.. 2026; 16(5): 2746-2756. doi:10.5455/OVJ.2026.v16.i5.17 Vancouver/ICMJE Style Akmam Ali Habeeb. Seed-extracted ZnO nanoparticles from Moringa peregrina to suppress aphid (Aphis craccivora) infestation on okra (Abelmoschus esculentus) using ace and COI RT-qPCR markers. Open Vet. J.. (2026), [cited June 26, 2026]; 16(5): 2746-2756. doi:10.5455/OVJ.2026.v16.i5.17 Harvard Style Akmam Ali Habeeb (2026) Seed-extracted ZnO nanoparticles from Moringa peregrina to suppress aphid (Aphis craccivora) infestation on okra (Abelmoschus esculentus) using ace and COI RT-qPCR markers. Open Vet. J., 16 (5), 2746-2756. doi:10.5455/OVJ.2026.v16.i5.17 Turabian Style Akmam Ali Habeeb. 2026. Seed-extracted ZnO nanoparticles from Moringa peregrina to suppress aphid (Aphis craccivora) infestation on okra (Abelmoschus esculentus) using ace and COI RT-qPCR markers. Open Veterinary Journal, 16 (5), 2746-2756. doi:10.5455/OVJ.2026.v16.i5.17 Chicago Style Akmam Ali Habeeb. "Seed-extracted ZnO nanoparticles from Moringa peregrina to suppress aphid (Aphis craccivora) infestation on okra (Abelmoschus esculentus) using ace and COI RT-qPCR markers." Open Veterinary Journal 16 (2026), 2746-2756. doi:10.5455/OVJ.2026.v16.i5.17 MLA (The Modern Language Association) Style Akmam Ali Habeeb. "Seed-extracted ZnO nanoparticles from Moringa peregrina to suppress aphid (Aphis craccivora) infestation on okra (Abelmoschus esculentus) using ace and COI RT-qPCR markers." Open Veterinary Journal 16.5 (2026), 2746-2756. Print. doi:10.5455/OVJ.2026.v16.i5.17 APA (American Psychological Association) Style Akmam Ali Habeeb (2026) Seed-extracted ZnO nanoparticles from Moringa peregrina to suppress aphid (Aphis craccivora) infestation on okra (Abelmoschus esculentus) using ace and COI RT-qPCR markers. Open Veterinary Journal, 16 (5), 2746-2756. doi:10.5455/OVJ.2026.v16.i5.17 |

