




| Research Article | ||
Open Vet. J.. 2026; 16(5): 2766-2780 Open Veterinary Journal, (2026), Vol. 16(5): 2766-2780 Research Article Determinants of genetic diversity, multidrug resistance, and virulence of Pseudomonas aeruginosa isolated from aquaculture fish in Central IraqMajida Malik Meteab Alshammari1, Shahad Ali Kamil1, Huda Hassan Gaber2, Orooba Meteab Faja2*, Zahira A. AL-Zuhairi2 and Balsam Miri Mizher Al Muhana31Department of Medical Microbiology, College of Medicine, Jabir Ibn Hayyan University of Medical and Pharmaceutical Sciences, Kufa, Iraq 2Department of Public Health, College of Veterinary Medicine, University of Al-Qadisiyah, Al-Diwaniyah, Iraq 3Department of Veterinary Microbiology, College of Veterinary Medicine, University of Al-Qadisiyah, Al-Diwaniyah, Iraq *Corresponding Author: Orooba Meteab Faja. Department of Public Health, College of Veterinary Medicine, University of Al-Qadisiyah, Al-Diwaniyah, Iraq. Email: orooba.faja [at] qu.edu.iq Submitted: 28/11/2025 Revised: 25/03/2026 Accepted: 07/04/2026 Published: 31/05/2026 © 2025 Open Veterinary Journal
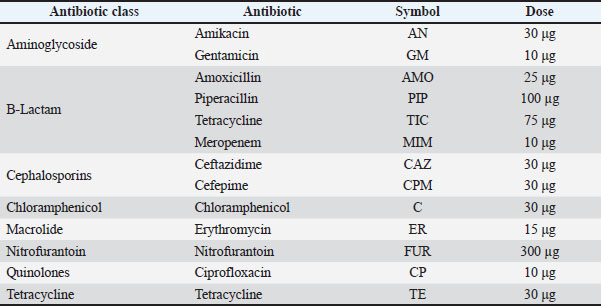
AbstractBackground: Pseudomonas aeruginosa is an opportunistic pathogen with the characteristics of a multidrug-resistant (MDR) antibiotic-resistant and highly virulent pathogen. Its presence in aquaculture systems poses significant threats and concerns for fish health, farm productivity, and potential zoonotic risk. Aim: This study aimed to assess the distribution of P. aeruginosa virulence genes, their antibiotic resistance profiles, and the genetic relationships of isolates from fish collected from major aquaculture systems in Central Iraq. Methods: A total of 158 fish were collected from six aquaculture sites in Central Iraq (three lake farms and three floating cage systems). Isolation was performed on cetrimide agar, and the species was identified through 16S rRNA amplification and sequencing. Antimicrobial susceptibility was tested using the disk diffusion technique for 13 antimicrobial agents in accordance with Clinical and Laboratory Standards Institute (CLSI) standards. The presence of nine virulence genes and five major antibiotic resistance genes was determined using polymerase chain reaction (PCR). To investigate clonal similarities/relations and geographic cladding, RAPD-PCR was performed. Results: Subsequently, 62 samples were examined and confirmed to be P. aeruginosa using 16S rRNA PCR and sequencing. Isolate phenotypic antimicrobial susceptibility testing showed 100% resistance to tetracycline and Cefuroxime / Cefuroxime sodium. Additionally, 92% of the samples were affected by MDR. Gene studies revealed that 41.9% of isolates contained all nine tested virulence factors. There were also distinct genetic groups in the RAPD-PCR analysis, possibly due to farming location. Pseudomonas aeruginosa was confirmed in 39.24% (62/158) of the samples. All isolates showed multiple antibiotic resistance indices of >0.2. The most frequently identified favorable attributes (virulence) of the pathogens were: fliC (74.19%), phz (70.96%), rhlI (61.29%), and pvd (56.45%). The formation of nine virulence patterns was recorded, with Pattern V1 (containing all genes) being the most prevalent (21%). The resistance genes aac(6′)-Ib, tetA, and blaTEM were found in 91.93%, 83.87%, and 82.25% of the isolates, respectively. RAPD-PCR revealed six geographically distinct clades that corresponded to particular fish farms, suggesting local clonal circulation and little intra-farm exchange. Conclusion: The results of the investigations confirm that P. aeruginosa inhabiting and genetically diverse populations in Iraqi aquaculture carry MDR and significantly virulent genotypes. Keywords: Antibiotic resistance, Aquaculture, Pseudomonas aeruginosa, Virulence genes. IntroductionThe global aquaculture market has been expanding rapidly over the past few decades, although the market has been experiencing declining growth rates and large bacterial pathogens affecting fish health and overall productivity. The global fauna of fish aquaculture suffers economically due to the presence of antibiotic multiresistant gram-negative Pseudomonas aeruginosa, which is suspected to be a pathogen to fish and zoonotically transmittable. The widespread use of antimicrobials and the selection of resistant strains increase the virulence of pathogens and their transfer to surrounding environments and even other ecosystems (Al-Shamari and Al-Khteeb, 2016; Safika et al., 2022a,b). The survival extended to the environment is made easier by the presence of certain outside genomic elements (Recchia and Hall, 1995). The need to evaluate the potential pathogens for disease control and the risks of these pathogens to humans and other higher-order organisms has grown rapidly. The presence of high levels of virulence and pathogenic potential has increased the need for understanding the circulating P. aeruginosa (within the aquaculture environment and ecosystem). The wide range of factors that contribute to the virulence and pathogenicity of P. aeruginosa can be attributed to multiple virulence factors, such as elastases, exotoxins, secretion systems, pigments, siderophores, and surface structures that aid in the adhesion and biofilm formation of the bacteria. The virulence factors of P. aeruginosa are also regulated by specific mechanisms of cell-to-cell communications, also known as quorum-sensing systems, that manage and control the actions of the bacteria in an infection. Recent molecular-level studies have shown that the virulence gene composition is not uniform in isolates from different ecological environments, indicating that the circulation of different virulent clones is shaped by specific environmental conditions (Brisse et al., 2009a,b; Al-Shuwaikh et al., 2021). Circulation of different virulent clones may also be associated with advanced tissue invasion, immunity evasion, and enhanced persistence in aquatic hosts (Rawya, 2008; Siu et al., 2011a,b). Pseudomonas aeruginosa has several virulence factors, such as exotoxins and specialized secretion systems. These include exotoxin A, the elastases LasA and LasB, alkaline protease, pyocyanin, and Type III secretion system (T3SS) ExoS, ExoT, ExoU, and ExoY. Other factors include the Type II secretion system and Type VI secretion system (T6SS). All of these factors together can lead to host tissue damage, immune response evasion, and disease progression. The presence of these systems has been described in clinically multidrug resistant (MDR) strains, increasing the concern about the potential for zoonotic transfer and environmental spread of these strains (Muhaidi et al., 2018a,b; Al-Anssari and Al-Charrakh, 2022). The rise of antimicrobial resistance on a global scale has conducted surveillance studies that combine the phenotype and genotype characterization of P. aeruginosa from aquaculture sources even more critical. Aminoglycoside, β-lactam, and tetracycline resistance determinants, such as aac(6′)-Ib, blaTEM, and tetA, have become more common in aquatic environments where antimicrobial residues are present (Michalska et al., 2014a,b,c; Saleh and Naji, 2022a,b,c). In addition, plasmid-mediated quinolone resistance genes, such as qnrS, cross bacterial populations and increase the likelihood of horizontal transfer to strains associated with humans (Robicsek et al., 2005; Park et al., 2006a,b). In Iraq, where aquaculture production has rapidly expanded, there have been increasing reports of antibiotic misuse and environmental contamination; however, little molecular data on P. aeruginosa remains. Thus, a detailed characterization of the prevalence, virulence determinants, resistance genes, and genetic relationship/relatedness of the isolates is essential for understanding the epidemiological situation of P. aeruginosa in aquaculture in Iraq and for designing relevant interventions. Pseudomonas aeruginosa is one of the most frequently encountered opportunistic bacterial pathogens in freshwater aquaculture systems, especially in settings with high levels of cultivation and frequent use of antimicrobials. Research in the Middle East and surrounding areas has reported the disease-related isolation of the bacteria in the range of 20%–45% and has recognized the significance of the pathogen in that region. Although there are some research results, there is still a scarcity of information that covers the profiles of antimicrobial resistance, distribution of virulence genes, and molecular characterization of P. aeruginosa from aquaculture in Iraq. The majority of available research has relied on phenotypic characterization without correlating resistance with virulence and clonal relationships. To the best of our knowledge, this is the first phenotypic, molecular, and genotypic characterization of P. aeruginosa isolates obtained from several aquaculture farms in Central Iraq. The present study was conducted to assess the distribution of P. aeruginosa virulence genes, its antibiotic resistance profiles, and the genetic relationships of the isolates from fish collected from major aquaculture systems in Central Iraq. Materials and MethodsSample collection and bacterial isolationFor a period of 3 months (July–October 2024), a total of 158 freshwater fish (Oreochromis niloticus; Cyprinus carpio) were collected aseptically from six different aquaculture sites in central Iraq (three land-based lake farms, three floating cage systems). The fish were placed in sterile, insulated cold containers to control the temperature and to avoid bacterial growth during transportation to the laboratory. At the laboratory, fish were individually dissected using flame-sterilized tools, and the gills, skin, and intestines were exposed and cut aseptically and individually placed on cetrimide agar (HiMedia, India, Cat. No. M023), and then aerobically incubated at 37°C for 24–48 hours. From these, the colonies were greenish blue and produced pyocyanin and were selected to ensure purity by subculturing on tryptic soy agar (HiMedia, India, Cat. No. M290). The isolates were presumed to be P. aeruginosa based on the green pigmentation, grape-like odor production, and oxidase positivity, along with growth on selective media. Altogether, 85 were presumed to be isolates, and from these pure subcultures were prepared for future work in the form of 20% glycerol stocks, which were made with Glycerol ≥99.5% (Sigma-Aldrich, USA; Cat. No. G5516) and kept at −80℃ for further use. Quality control from Tryptic Soy Agar (TSA) cultured cellsThe GeneJET Genomic DNA Purification Kit was used (Thermo Fisher Scientific, USA, Cat. No. K0721) to obtain Genomic DNA from TSA overnight cultures following the supplier's guide, producing DNA of sufficient quality for polymerase chain reaction (PCR). Identification of P. aeruginosa was confirmed using the PCR to amplify the 16S rRNA gene with universal primers27F (5′-AGAGTTTGATCATGGCTCAG-3′) as forward and 1492R (5′-GGTTACCTTGTTACGACTT-3′) as reverse primer. In a total of 50 µl, 25 µl of DreamTaq Green PCR Master Mix (2X) (Thermo Fisher Scientific, USA; Cat. No. K1081), 0.4 µM of primers for each (Integrated DNA Technologies, USA), 10 to 100 ng of the genomic DNA template, and the appropriate volume of nuclease free water (Thermo Fisher Scientific, Cat. No. R0581). Amplification used the cycling conditions described in the reference protocol in an Eppendorf Mastercycler® Nexus Gradient (Eppendorf, Germany): an initial 94°C denaturation for 2 minutes; 94°C for 1 minute, 55°C for 1.5 minutes, 72°C for 1 minute (35 times cycle), and for 3 minutes a final 72°C extension in addition to the previously mentioned conditions. PCR products were extracted from the gel and viewed for the gel’s DNA Ethidium Bromide. The gel (1%) was manufactured as UltraPure™ (Invitrogen, USA, Cat. No. 16500-500). Based on the distinct antimicrobial profiles, virulence factor gene patterns, and place of farm origin, 10 representative isolates were selected for the 16S rRNA gene sequencing. This was to ensure the coverage of the documented morphological, and spatial distribution of the diversity from the 62 confirmed isolates. E7637 using a UV transilluminator (Bio-Rad, USA). The PCR products were purified using the GeneJET PCR Purification Kit (Cat. No. K0701) and sent for Sanger sequencing (Macrogen Inc., South Korea). The sequenced 16S rRNA gene fragments were compared with the National Center for Biotechnology Information (NCBI) nucleotide (nt) database using the BLASTn algorithm. The parameters were kept as the default, and included a word size of 28, match/mismatch scores of +1/−2, penalties of 5 and 2 for gap opening and extension, respectively, and the low-complexity filter turned on. Pseudomonas aeruginosa sequences in GenBank were used as the standard reference for identification, and the identification was made based on sequence similarity of 98.7%. Only alignments with an almost zero E-value that scored the highest were used for identification. Antimicrobial susceptibility testSusceptibility of all 62 confirmed P. aeruginosa isolates were analyzed employing the Kirby-Bauer disk diffusion technique that complies with Clinical and Laboratory Standards Institute (CLSI) MM18 (2022). Very bacterial suspensions were prepared using sterile saline, matching a zero-point five McFarland standard, which was calibrated using a DEN-1 McFarland Densitometer. Each suspension was spread evenly onto Mueller-Hinton Agar, and antibiotic disks were safely placed. Employing the following antibiotics: 30 µg Amikacin, 10 µg Gentamicin, 25 µg Amoxicillin, 100 µg Piperacillin, 75 µg Ticarcillin, 10 µg Meropenem, 30 µg Ceftazidime, 30 µg Cefepime, 30 µg Chloramphenicol, 15 µg Erythromycin, 300 µg Nitrofurantoin, 10 µg Ciprofloxacin, and 30 µg Tetracycline. The plates were kept in incubators at 37 and diagnosed for 18–24 hours. The multiple antibiotic resistance (MAR) index was calculated for each isolate as the ratio of the number of antibiotics to which the isolate was resistant divided by the total number of antibiotics tested. Isolates were grouped into nine antibiogram patterns (P1–P9) based on shared resistance profiles across the tested antibiotics, with higher MAR values indicating exposure to environments with elevated antimicrobial selective pressure. Although erythromycin and chloramphenicol were considered to have minimal clinical efficacy against P. aeruginosa, these agents were included to determine the intrinsic patterns of resistance and allow comparison to previous regional surveillance studies of environmental isolates. Interpretation and P. aeruginosa ATCC 27853 acted as the quality control strain. The CLSI breakpoints were used (Table 1). Table 1. Antibiotics used in this study.
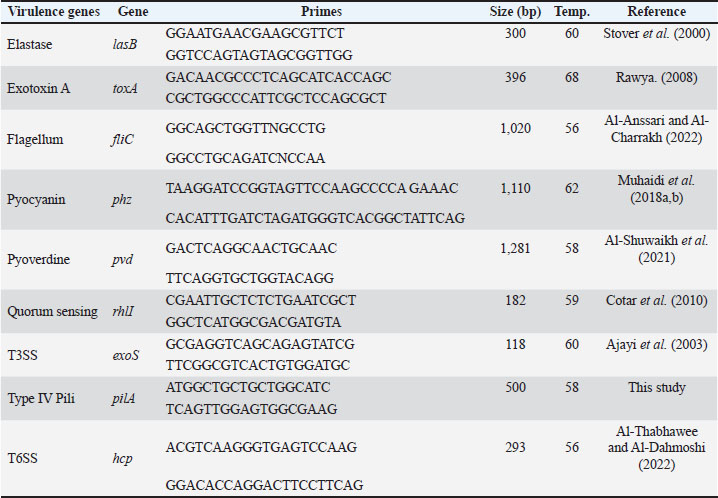
Detection of virulence genes via PCRUsing standard PCR, we screened for nine major virulence genes, including lasB, toxA, fliC, phz, pvd, rhlI, exoS, pilA, and hcp, for which we detail primer sequences, amplicon sizes, annealing temperatures, and references in Table 2. The PCR reaction mixtures consisted of 25 µl of EconoTaq® PLUS GREEN 2X Master Mix (Lucigen, USA; Cat. No. 30033-2), 1 µl of each primer at a concentration of 10 pmol/µl (Macrogen, Korea), 1 µl of the template DNA, and DNAse-free water to a final volume of 50 µl. The PCR products were loaded into 1.5% agarose gels (Invitrogen), running in 1X TAE buffer (Thermo Fisher; Cat. No. B49) and were stained with ethidium bromide. We used a BioRadGelDoc XR+ for imaging and performing documentation and analysis to capture the images. Table 2. Primers for detecting virulence genes.
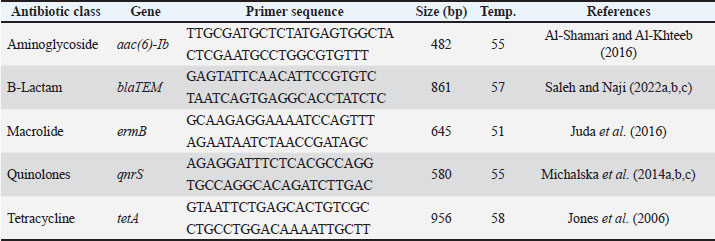
Identification of antibiotic resistance genesThe aac(6′)-Ib, blaTEM, ermB, qnrS, and tetA genes were assessed for PCR amplification using gene-specific primers. To conduct PCR amplification, DreamTAq Master Mix (Cat. No. K1081) was used in a reaction volume of 50 µl which contained 25 µl of Master Mix, 10 pmol of the primers, and approximately 50 ng of template DNA (Table 3). Amplicons were resolved on 1.5% agarose gels, and a 100 bp DNA ladder (Cat. No. 15628-019) was used for comparison. Molecular confirmation with 16S rRNA gene sequencing was performed for the 10 representative P. aeruginosa isolates (PF02, PF05, PF12, PF19, PF24, PF28, PF37, PF43, PF49, and PF59), for which distinct patterns of antibiotic susceptibility, virulence genes, and geography were used in the previous selection of the isolates. Sequence identity was confirmed using the BLASTn tool with the NCBI database for nucleotides, and all confirmed isolates were assigned accession numbers. This selection procedure and molecular characterization were the same as those used in previous studies for P. aeruginosa isolates from aquaculture fish in Central Iraq, where the ecological patterns of this pathogen were documented in terms of its genetic diversity, virulence, and antimicrobial resistance. Table 3. Primers for the detection of antibiotic resistance genes.
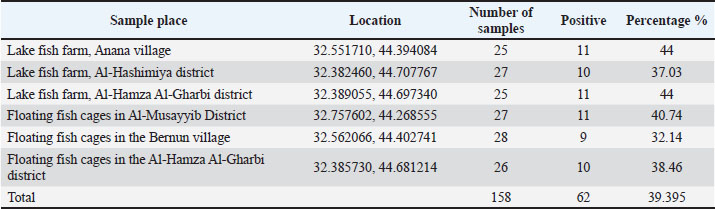
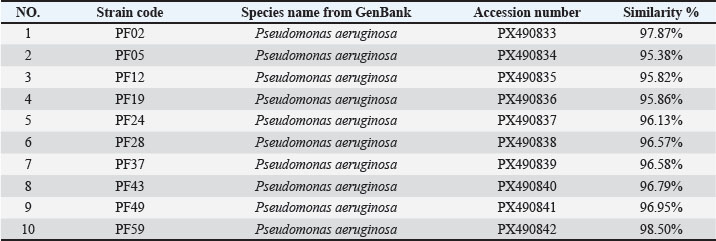
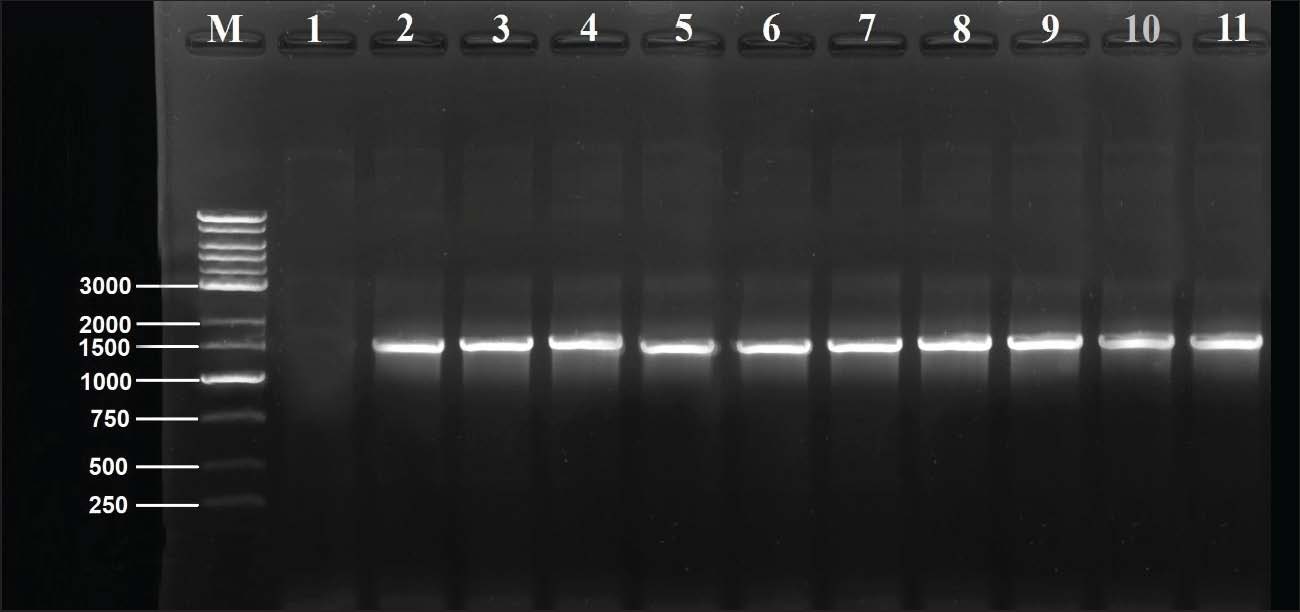
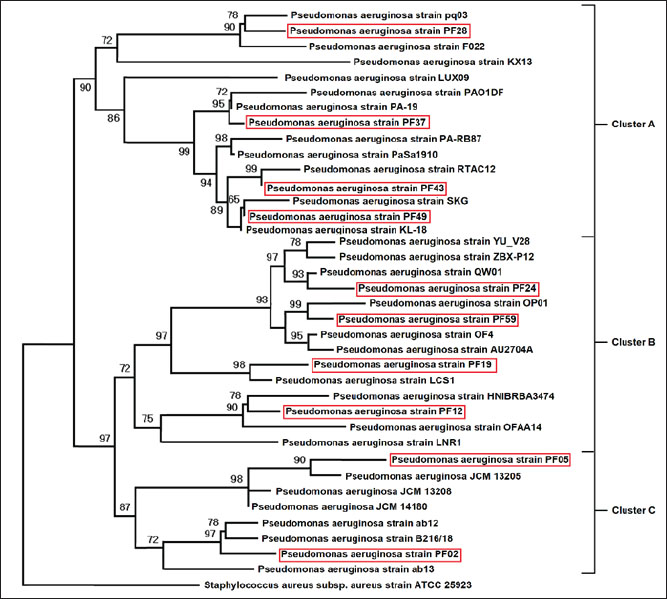
Resistance genes were used to create gene pattern profiles (P1–P6) to describe the different combinations of detected resistance genes. RAPD-PCR molecular typingThe OPA-09 primer (5'-GGGTAACGCC-3') from Macrogen (Korea) was used to assess the genetic distance between the isolates using RAPD-PCR. For the amplifications, 50-μl reactions (25 μl EconoTaq® PULS GREEN 2X Master Mix (Lucigen, USA), 1 μl primer, 1 μl template DNA), and 23 μl of nuclease-free water were prepared. The cycling conditions were as follows: initial denaturation at 94°C for 5 minutes, followed by 45 cycles of 94°C for 1 minute, 35°C for 1 minute, and 72°C for 2 minutes. The final extension was conducted at 72°C for 7 minutes. The PCR amplification products were then run on 1.5% agarose gel for 90 minutes at 80 V. Images of the gel were obtained and analyzed using ImageJ v1.53. Dendrograms were constructed using PHYLIP v3.697 with the Unweighted Pair Group Method with Arithmetic Mean (UPGMA) method and the Pearson correlation coefficient. These methods were used to assign clonal groups at a distance of 80% of RAPD PCR similarity. Statistical analysesStatistical analyses were conducted using the IBM SPSS Statistics software. The quantitative variables were the diameters of the zones of antimicrobial inhibition and the molecular expression values, which were summarized as mean ± standard deviation. Before performing any inferential analysis, data were examined for normality using the Shapiro-Wilk test, and Levene’s test was used to assess the equality of variances. Since these assumptions were met, we conducted parametric tests. For comparisons of more than two independent groups, one-way analysis of variance was conducted, and Tukey’s post hoc test was used to adjust for multiple pairwise comparisons. For the variables of the categories, which were the distribution of the resistance phenotypes and the frequencies of the isolates, the chi-square test was used where appropriate. A p-value of 0.05 was considered statistically significant. The sampling frame reflected the exploratory nature of the study rather than being hypothesis-driven and was therefore determined by the total number of non-duplicate isolates collected during the defined sampling period. Therefore, an a priori power analysis was not conducted, which you have acknowledged as a limitation to avoid overgeneralization, and the statistical results were interpreted with due care. As you have noted, the statistical techniques utilized were appropriate and in keeping with accepted biostatistical methods in the field of microbiology and antimicrobial susceptibility testing (George and Mallery, 2019). Ethical approvalThe Scientific and Ethical Committee, College of Veterinary Medicine, University of Al-Qadisiyah approved this study (Approval number 4914, 14/11/2024). ResultsFindings of molecular identification and 16S rRNA sequencingMolecular confirmation was performed using conventional PCR with the 16S rRNA gene for 10 representative isolates (PF02, PF05, PF12, PF19, PF24, PF28, PF37, PF43, PF49, and PF59). Electrophoresis of PCR products confirmed that all isolates were expected species P. aeruginosa (Table 4), as all amplified the targeted 1,500 bp region (Fig. 1). Sanger sequencing of the amplified products, followed by comparison using the BLAST tool, indicated that the sequenced products were 95.38%–98.50% similar to other P. aeruginosa sequences in GenBank, with all isolates forming closed clades to the respective reference sequences. Table 5 shows the similarity percentages and accession numbers for the submitted P. aeruginosa sequences to GenBank. The isolates were also submitted to GenBank as part of a phylogenetic study, which showed that the isolates belong to distinct clades (A, B, C) with high bootstrap support and a unique alignment to the P. aeruginosa lineages (Fig. 2). Table 4. Prevalence of P. aeruginosa infection in fish from different aquaculture locations.
Table 5. BLAST similarity analysis of 16S rRNA sequences of isolates with GenBank records.
Fig. 1. Agarose gel (1%) electrophoresis of 16S rRNA PCR products (~1,500 bp) of selected P. aeruginosa isolates. Lane M: 1kb ladder marker; Lane 1: negative control; Lane 2: PF12; Lane 3: PF19; Lane 4: PF43; Lane 5: PF24; Lane 6: PF28; Lane 7: PF05; Lane 8: PF49; Lane 9: PF59; Lane 9: PF37; Lane 10: PF02.
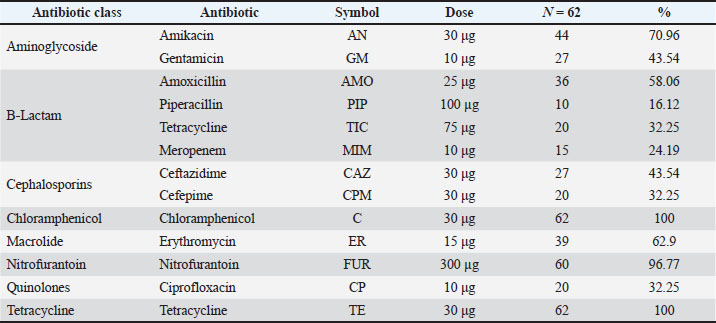
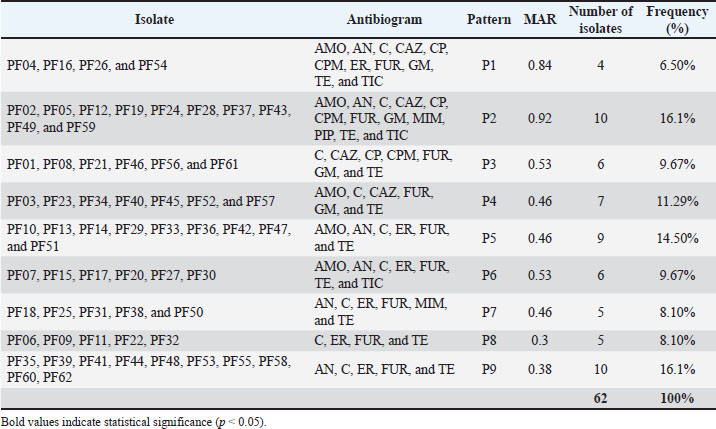
Fig. 2. Phylogenetic tree of 16S rRNA sequences showing evolutionary relationships among isolates. The 16S rRNA gene sequences were aligned using the ClustalW method in MEGA software (version 11). Phylogenetic relationships were constructed using the neighbor-joining method with the Kimura two-parameter model. Only bootstrap values over 50 were displayed, and the tree topology robustness was tested with 1,000 bootstrap repetitions. Reference sequences of P. aeruginosa obtained from GenBank were added for comparative analysis. Findings of profiles of resistance antibioticsRegarding the 62 bacterial isolates, we observed a striking degree of antimicrobial resistance (AMR), with total resistance for the tested antibiotics at 100% for both chloramphenicol and tetracycline. The Nitrofurantoin resistance was virtually complete at 96.77%. Other drugs with significant resistance included amikacin (70.96%), erythromycin (62.9%), amoxicillin (58.06%), and ceftazidime (43.54%). Resistance was statistically lower with piperacillin at 16.12%, meropenem at 24.19%, and ciprofloxacin at 32.25%, all of which indicated concern levels of resistance (p < 0.001). All isolates had a minimum inhibitory concentration of antibiotics resistance greater than 0.2 µg/ml, indicating that they had all been in severely antimicrobial-resistant environments. Antibiotic resistance levels in percentage are shown in Table 6. Nine distinct antibiogram patterns (P1–P9) were identified, with patterns P2 and P9 as the most common (16.1%, respectively), with MAR values of 0.30 and 0.92 (Table 7). Table 6. Antibiotic resistance profiles of P. aeruginosa isolates.
Table 7. The antibiotic resistance patterns (P1–P9) and MAR indices of the isolates.
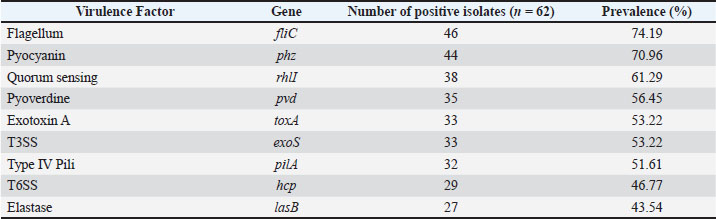
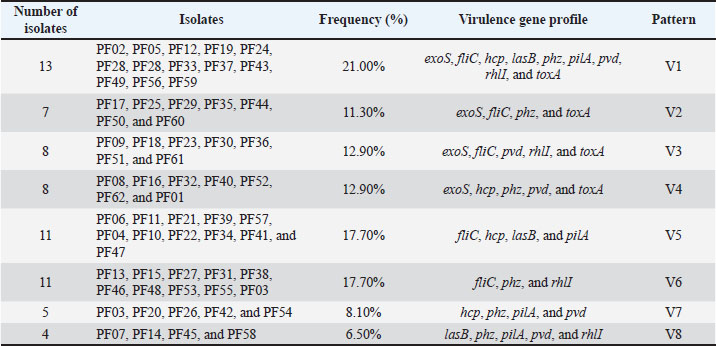
The geographical and genetic distribution of virulence genes across the different isolate synonyms of virulence PCR of the nine most important virulence factors provided preliminary evidence of the geographical and genetic Hirbodel scope of virulence determining factors of the isolates. Most of the isolates were positive for fliC (74.19%) and phz (70.96%), as well as for motility and phenazine production. The other positively amplified genes were rhlI (61.29%), pvd (56.45%), and for exoS (52.48%), and pilA (51.61%), demonstrating that most isolates possessed different virulence traits of adhesion for an iron and toxin of secretion of a quorum. Significantly lower frequencies were noted for hcp (46.77%) and lasB (43.54%). Table 8 shows the overall virulence and gene frequencies. On the basis of various analyzed virulence genes, the virulence patterns were grouped into eight clades V1–V8, of which V1 was the most virulent, with a dominance of all nine virulence genes in the clade (21.0%). Table 9 shows the distribution of the gene amplification patterns of virulence, and the patterns of virulence amplification genes are also shown in Figure 3. Table 8. Prevalence of virulence genes among P. aeruginosa isolates.
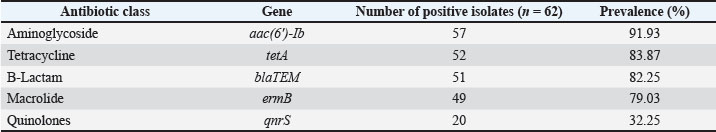
Table 9. Distribution of V1–V8 virulence gene patterns among isolates.
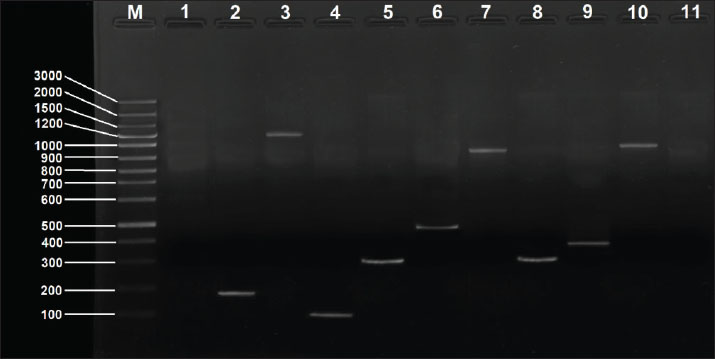
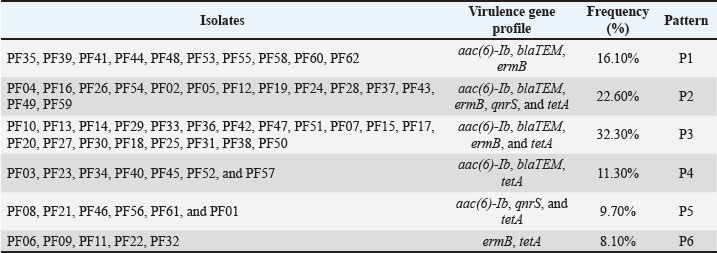
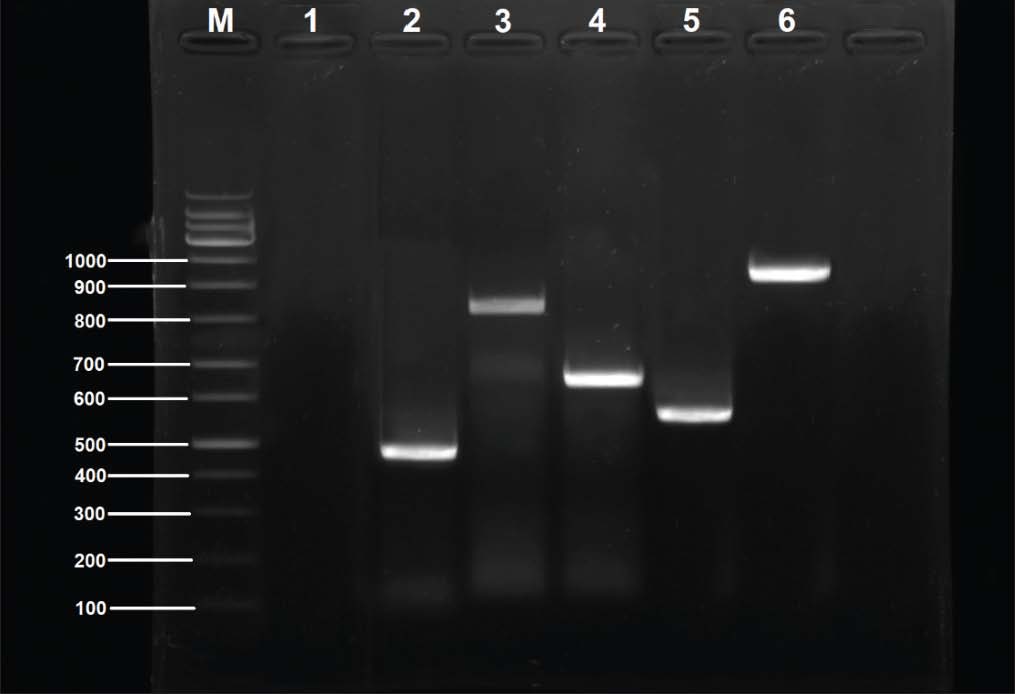
Fig. 3. PCR-agarose gel (1.5%) amplification patterns of virulence genes detected in the P. aeruginosa PF24 isolate. Lane M: 100-bp DNA ladder; Lane 1: negative control; Lane 2: rhlI gene; Lane 3: pvd gene; Lane 4: exoS gene; Lane 5: lasB gene. Lane 6: pilA gene. Lane 7: phz gene Lane 8: hcp gene; Lane 9: toxA gene; Lane 10: fliC gene. Findings of resistance genesScreening for five resistance determinants revealed a high prevalence of genes responsible for resistance to aminoglycosides, β-lactams, macrolides, and tetracyclines. The prevalence of the aac(6′)-Ib gene was the highest (91.93%), followed closely by tetA (83.87%, blaTEM at 82.25%), and ermB (79.03%). The plasmid-mediated qnrS gene was present in 32.25% of the isolates. A strong correlation was found between the prevalence of the genes and the resistance phenotypes recorded in the earlier mentioned study. The prevalence of each gene is shown in Table 10. Using a combination of genotypes, six resistance gene patterns (P1–P6) were described. P3 was the most common (32.3%) and P2 (22.6%), the only pattern containing all five genes. The patterns are summarized in Table 11, and representative results of the PCR confirmations are shown in Figure 4. Table 10. Distribution of antibiotic resistance genes among P. aeruginosa isolates.
Table 11. Distribution of the antibiotic resistance gene patterns (P1–P6).
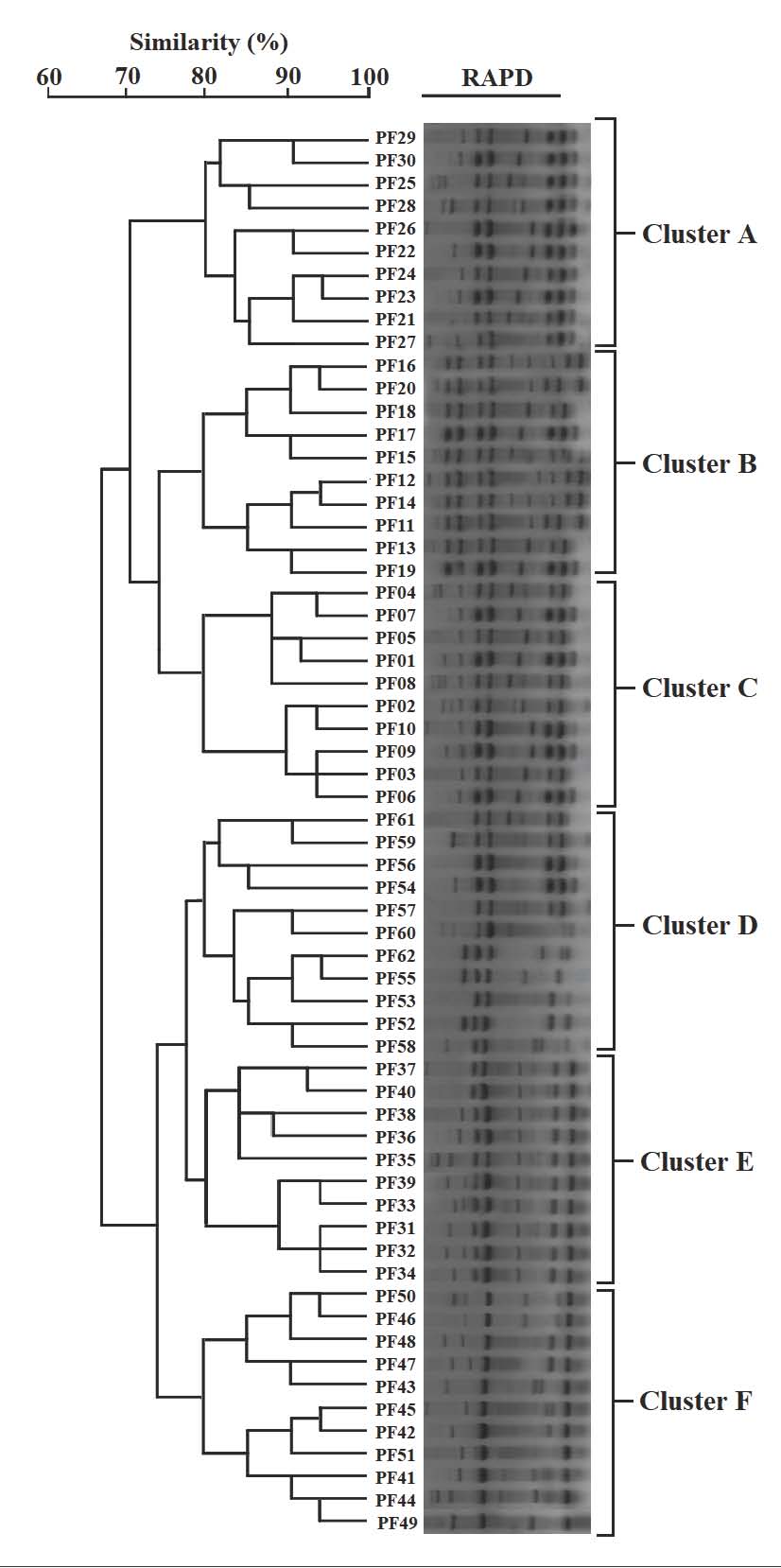
Fig. 4. Amplification of antibiotic resistance genes detected in P. aeruginosa PF02 isolate by PCR agarose gel (1.5%). Lane M: 100 bp DNA ladder. Lane 1: Negative control. Lane 2: aac(6)-Ib gene. Lane 3: blaTEM gene. Lane 4: ermB Lane 5: qnrS gene Lane 6: tetA gene. Findings of RAPD-PCR molecular typing and genetic relatednessWhile conducting RAPD-PCR analysis for the primer OPA-09, notable genetic variability was observed among the 62 isolates, as distinct but reproducible banding patterns were constructed for dendrogram analysis. The UPGMA clading determined six major clades (A–F) at an 80% similarity threshold. Each clade was directly correlated to its geographical location: clades A–C comprised isolates from lake-based farms, while clades D–F incorporated isolates from floating cage systems. This geographical configuration indicates a localized clonal amplification with little to no interfarm movement. The dendrogram illustrating these relationships is shown in Figure 5, with notable segregation of isolates from different aquaculture systems showing the molecular epidemiological difference among the various sites.
Fig. 5. RAPD-PCR dendrogram illustrating the genetic relatedness of P. aeruginosa isolates. DiscussionPseudomonas aeruginosa is an opportunistic pathogen in aquatic farming and has been regularly documented in the collection of fish, water, and sediment samples in both fresh and marine water systems. It occurs in aquaculture because of off-balanced farming systems, excessive organic waste, and poor water quality. A number of research articles document that P. aeruginosa has a high degree of both intrinsic and acquired antimicrobial resistance to aquaculture, particularly to beta-lactams, tetracyclines, and aminoglycosides. Resistance to the aforementioned antibiotics is due to little to no outer membrane porin, an overexpression of efflux pumps, and the high incidence of the acquisition and preservation of resistance genes through lateral genetic transduction. Consequently, P. aeruginosa has been a persistent species for a long time, and it has been very hard to restrict the spread of this species in aquaculture, especially in the farming of fish, which raises the level of concern for the health of aquaculture fish, the environment in which the resistance genes spread, and the health of the people. The detection of P. aeruginosa in 39.24% of the tested fish shows that the bacterium is somewhat widespread in the reported cases of problems in certain aquaculture systems. However, this prevalence can only be interpreted as relative because it pertains only to the farms that were sampled and does not indicate the situation in other aquaculture systems. This result confirms observations made about the bacterium as a high-frequency occurring pathogen that has the ability to inhabit various freshwater ecosystems that are rich in nutrients, along with the presence of antimicrobials in the water (Kim et al., 2005; Safika et al., 2022a,b). The prevalence of P. aeruginosa from poultry and poultry intensively farmed animals (broilers) reported by Safika et al. (2022a,b) is 39.24%, which is similar to that reported by Safika et al. (2022a,b) and lower than the figures reported in various clinical and environmental research works in Europe, which is characterized by more structured hygiene practices, which in turn limit contamination of the environment (Michalska et al., 2014a,b,c). The fact that several phylogenetic clades correlate positively with specific geographic locations indicates that there is little to no exchange of pathogens among farms, and there is a possibility of clonal evolution in the semi-isolated aquaculture systems, which depict locality described in virulent P. aeruginosa lineages in different ecosystems of the world (Brisse et al., 2009a,b; Siu et al., 2011a,b). In contrast to the clinical strains that are highly recombinant, the unique cladding that is evident in the current study suggests more of a local adaptation that is not accompanied by a high degree of horizontal gene transfer. Overall, P. aeruginosa isolates showed no resistance to chloramphenicol and tetracycline. Although some resistance genes were identified through PCR, they were not as common as expected. Such discrepancies between gene and phenotypic responses are not surprising and are expected due to the intrinsic mechanisms of antimicrobial resistance exhibited by P. aeruginosa. Resistance to chloramphenicol and tetracycline in P. aeruginosa is often attributed to the organism’s inherent mechanisms. Overexpression of certain multidrug efflux pumps, such as MexAB-OprM and MexXY-OprM, and the ability to extrude multiple classes of antibiotics would phenotypically typify resistance without the need for specific acquired genes. In P. aeruginosa, resistance mechanisms may include reduced outer membrane permeability, some regulatory mutations concerning the expression of the efflux pump, or some chromosomally encoded factors that are infrequently subject to routine conventional PCR. In this study, resistance screening via PCR was restricted to some notable resistance genes; thus, all documented genetic mechanisms related to resistance to tetracycline or chloramphenicol were not evaluated. Therefore, just because some genes were not found via PCR, other mechanisms of resistance are not available. Such facts were the limitation that genotype–phenotype correlations must address and highlight the need for coupling phenotypic susceptibility testing with molecular methods to gain a deeper insight into the mechanisms driving antimicrobial resistance of P. aeruginosa. These results echo the findings of studies focusing on agricultural cross-resistance or on the correlation between food animal resistance and rampant antimicrobial overuse (Chen et al., 2004; Kim et al., 2005). The very high prevalence of the resistance genes aac(6’)–Ib, blaTEM, ermB, and tetA parallels that found in clinically-sourced specimens from the Middle East and other countries in Asia (Al-Shamari and Al-Khteeb, 2016; Saleh and Naji, 2022a,b,c) and suggests that aquaculture systems should be viewed as contributing to a wider differential AMR burden in the region. The relatively lower prevalence of qnrS in our isolates is not in accordance with the most documented high prevalence of qnrS in U.S. hospital strains (Robicsek et al., 2005; Park et al., 2006a,b), and it may be an indication of the pattern of quinolone use in the country. The positive association between the phenotypic resistance and the genetic markers as established in this study reinforces the practical significance of these genes and parallels the findings in other instances clinically and agronomically where the aminoglycosides and β-lactam resistant genes became established (Michalska et al., 2014a,b,c; Safika et al., 2022a,b). The diversity and prevalence of fliC, phz, rhlI, and toxA indicate that Iraqi aquaculture isolates have a similar pathogenic potential to internationally reported virulent clones. These findings align with earlier studies that reported similar levels of virulence gene prevalence in isolates from burn patients, cases of urinary tract infection, and farm animals (Rawya, 2008; Muhaidi et al., 2018a,b; Al-Shuwaikh et al., 2021). Of the most concern, 21% of the isolates harbored complete virulence profiles of the nine-gene (V1). Such virulence has been associated with severe invasive infections in humans and animals (Brisse et al., 2009a,b; Siu et al., 2011a,b). In contrast to reports of the low prevalence of elastase or T6SS in environmental isolates, our study showed the widespread presence of these traits, indicating that the local fish farms may have conditions that favor the maintenance of such a highly virulent arsenal. The combination of high-virulence genotypes and a MDR phenotype supports the concept of “high-risk clones” and highlights the potential for these high-virulence genotypes to be zoonotically transmitted or spill over into human health. The zoonotic implications in this study are based on molecular comparison with other pathogenic strains and corresponding literature, rather than evidence from direct host–pathogen interactions, which are absent from this study. Most recent studies have stressed the importance of the biological relevance of the identified molecular determinants being supported by empirical validation of the molecular interactions and the stability of the molecular determinants, which connects the detection of genotypes to their functional activity, under physiological conditions. Detection of virulence-associated genes only provides a clue of theoretical pathogenicity and does not offer proof of a functioning pathogen, as no in vivo or phenotypic validation testing was performed. Pseudomonas aeruginosa genotypes, localized dissemination of, suggest partial association with farm location; distinct genetic clades were revealed through RAPD-PCR analysis. Some clades show resistance and virulence gene patterns, indicating that genetically related isolates have adaptive traits. However, the observed variability reflects the selective pressures and influence of horizontal gene transfer. RAPD-PCR analysis provides preliminary insight into genetic relatedness and suggests the need for higher order typing to confirm definitive lineage relationships. RAPD-PCR analysis revealed distinct genetic clades that were partially associated with farm location, suggesting that specific P. aeruginosa genotypes were locally distributed. The patterns of prevalence within the aquaculture systems sampled influence genotype distribution rather than the underlying mechanisms of genetic stabilization or environmental persistence, which are experimentally verified mechanisms that are absent in this study. Despite some Random Amplified Polymorphic DNA patterns clustering some isolates together, considering such patterning as evidence for the lack of cloning or restriction horizontal gene transfer would be erroneous, as P. aeruginosa is notorious for its mobile genetic elements and recombination events. While RAPD-PCR is a quick and inexpensive method for evaluating genetic diversity, it is less precise and offers lower resolution than whole-genome sequencing, multilocus sequence typing, and single nucleotide polymorphism-based phylogenetic analyses. Future research must combine molecular screening methods and functional validation strategies, especially intracellular assays, in vivo infection studies, and precise-resolution genomics, to gain a more complete understanding of the biological actions and durability of the identified resistance and virulence elements. Limitations of the studyThe current study was constrained by the lack of environmental sampling, phenotypic virulence assays, and high-resolution genotyping strategies. In addition, the levels of antibiotic residues in water or feed were not measured. Future studies should include these to clarify selective pressures and transmission patterns. Some details regarding the conclusions made about the infection models pertaining to virulence and resistance should be noted. These conclusions were based on molecular screening procedures, and the functional activity of the determinants was not validated through functional assays performed during direct intracellular infections. ConclusionThis study shows that Iraq has a large number of MDR microorganisms and a large number of virulent P. aeruginosa present in aquaculture systems. Resistance to a lot of key genes, as well as a lot of virulence factors, along with four different distinct clades of the organism, RAPD-PCR located in different areas, shows how the organism can remain, adapt, and move within the aquaculture fish farms under high sand core pressures. This also shows how vital it is to have better antibiotic control, along with a high biosecurity level. AcknowledgmentThe authors would like to express their gratitude to the College of Veterinary Medicine for providing laboratory facilities and technical support throughout the study. Conflict of interestThe authors declare that there are no conflicts of interest existed in this work.FundingThis research was self-funded by the authors with no external financial support. Authors’ contributionsAll authors have contributed to this study. Data availabilityData are available upon request from the corresponding author. ReferencesAjayi, T., Allmond, L.R., Sawa, T. and Wiener-Kronish, J.P. 2003. Single-nucleotide polymorphisms in Type III secretion toxin genes of Pseudomonas aeruginosa. J. Clin. Microbiol. 41(8), 3526–3531. Al-Anssari, M.J. and Al-Charrakh, A.H. 2022. Partial purification of type A and B flagellin from Pseudomonas aeruginosa for preclinical use. Int. J. Health. Sci. 3661, 3661–3670. Al-Shamari, R.K. and Al-Khteeb, S.N. 2016. Molecular characterization of aminoglycosides resistance in Pseudomonas aeruginosa. Iraqi. J. Sci. 1150, 1150–1157. Al-Shuwaikh, A.M.A., Al-Shwaikh, R.M.A. and Al-arnawtee, A.F. 2021. Study of pvdA gene sequencing from Pseudomonas aeruginosa isolated from burn infections. Iraqi. J. Agric. Sci 17(2), 1–12. Al-Thabhawee, M.H.J. and Al-Dahmoshi, H.M. 2022. Molecular investigation of outer membrane channel genes among MDR Pseudomonas aeruginosa isolates. Rep. Biochem. Mol. Biol. 11(1), 102–111. Brisse, S., Fevre, C., Passet, V., Issenhuth-Jeanjean, S., Tournebize, R., Diancourt, L. and Grimont, P. 2009a. Virulent clones of Pseudomonas aeruginosa: genomic and phenotypic characterization. PLos One 4(3), e4982. Brisse, S., Fevre, C., Passet, V., Issenhuth-Jeanjean, S., Tournebize, R., Diancourt, L. and Grimont, P. 2009b. Virulent clones of Pseudomonas aeruginosa: identification and evolutionary scenario based on genomic and phenotypic characterization. PLos One 4(3), 4982. Chen, S., Zhao, S., White, D.G., Schroeder, C.M., Lu, R., Yang, H., Mcdermott, P.F., Ayers, S. and Meng, J. 2004. Characterization of multiple-antimicrobial-resistant Salmonella serovars isolated from retail meats. Appl. Environ. Microbiol. 70(1), 1–7. Cotar, A.I., Chifiriuc, M.C., Dinu, S., Pelinescu, D., Banu, O. and Lazar, V. 2010. Influence of probiotic culture fractions on Pseudomonas aeruginosa quorum sensing gene expression. Roum. Arch. Microbiol. Immunol. 69(4), 213–223. George, D. and Mallery, P. 2019. IBM SPSS statistics 26 step by step: a simple guide and reference, Jones, C.H., Tuckman, M., Murphy, E. and Bradford, P.A. 2006. Identification of a tetracycline-resistance determinant homologue in Gram-negative rods. J. Bacteriol. 188(20), 7151–7164. Juda, M., Chudzik-Rzad, B. and Malm, A. 2016. MLSB resistance genotypes and susceptibility among staphylococci. Mem. Inst. Oswaldo. Cruz. 111, 155–160. Kim, S.H., Wei, C.I., Tzou, Y.M. and An, H. 2005. Multidrug-resistant Pseudomonas aeruginosa from farm environments and retail products. J. Food. Prot. 68(10), 2022–2029. Michalska, A.D., Sacha, P.T., Ojdana, D., Wieczorek, A. and Tryniszewska, E. 2014a. Aminoglycoside and fluoroquinolone resistance in Pseudomonas aeruginosa. Braz. J. Microbiol. 45, 1455–1458. Michalska, A.D., Sacha, P.T., Ojdana, D., Wieczorek, A. and Tryniszewska, E. 2014b. Prevalence of aminoglycoside and fluoroquinolone resistance in Pseudomonas aeruginosa. Braz. J. Microbiol. 45, 1455–1458. Michalska, A.D., Sacha, P.T., Ojdana, D., Wieczorek, A. and Tryniszewska, E. 2014c. Prevalence of resistance to aminoglycosides and fluoroquinolones among Pseudomonas aeruginosa strains in a university hospital in northeastern Poland. Braz. J. Microbiol. 45, 1455–1458. Muhaidi, M.J., Aziz, L.M. and Ahmed, M.N. 2018a. Genetic evaluation of phenazine synthesized by Pseudomonas aeruginosa isolated from genital tract of farm animals. Iraqi J. Agric. Sci. 49(2). Muhaidi, M.J., Aziz, L.M. and Ahmed, M.N. 2018b. Genetic evaluation of phenazine synthesized by Pseudomonas aeruginosa isolated from genital tracts of farm animals. Iraqi J. Agric. Sci. 49(2). Park, C.H., Robicsek, A., Jacoby, G.A., Sahm, D. and Hooper, D.C. 2006a. Prevalence in the United States of aac(6′)-Ib-cr encoding a ciprofloxacin-modifying enzyme. Antimicrob. Agents. Chemother. 50(11), 3953–3955. Park, C.H., Robicsek, A., Jacoby, G.A., Sahm, D. and Hooper, D.C. 2006b. Prevalence of aac(6′)-Ib-cr in U.S. isolates. Antimicrob. Agents. Chemother. 50(11), 3953–3955. Rawya, B. 2008. Pseudomonas aeruginosa exotoxin A as a virulence factor in burn wound infections. J. Clin. Microbiol 49(2), 151–157. Recchia, G.D. and Hall, R.M. 1995. Gene cassettes: a new class of mobile element. Microbiology 141, 3015–3027. Robicsek, A., Sahm, D.F., Strahilevitz, J., Jacoby, G.A. and Hooper, D.C. 2005. Plasmid-mediated quinolone resistance distribution in the U.S. Antimicrob. Agents Chemother. 49(7), 3001–3003. Safika, S., Nilasari, Z. and Pasaribu, F.H. 2022a. Antibiotic resistance genes in Pseudomonas aeruginosa from broiler chickens. J. Appl. Pharm. Sci. 12(7), 190–198. Safika, S., Nilasari, Z. and Pasaribu, F.H. 2022b. Detection of antibiotic resistance coding genes in Pseudomonas aeruginosa isolated from broiler chickens in West Java, Indonesia. J. Appl. Pharm. Sci. 12(7), 190–198. Saleh, M.A.A.J.M. and Naji, H.F. 2022a. Detection of blaTEM, blaCTX-M, and blaSHV genes in clinical isolates of multidrug-resistant Pseudomonas aeruginosa. Int. J. Health. Sci. 6(S7), 3239–3253. Saleh, M.A.A.J.M. and Naji, H.F. 2022b. Detection of β-lactamase genes in MDR Pseudomonas aeruginosa. Int. J. Health. Sci. 6(S7), 3239–3253. Saleh, M.A.A.J.M. and Naji, H.F. 2022c. Detection of β-lactamase genes in multidrug-resistant Pseudomonas aeruginosa. Int. J. Health. Sci. 6(S7), 3239–3253. Siu, L.K., Fung, C.P., Chang, F.Y., Lee, N., Yeh, K.M., Koh, T.H. and Ip, M. 2011a. Molecular typing and virulence analysis of Pseudomonas aeruginosa serotypes. J. Clin. Microbiol. 49(11), 3761–3765. Siu, L.K., Fung, C.P., Chang, F.Y., Lee, N., Yeh, K.M., Koh, T.H. and Ip, M. 2011b. Molecular typing and virulence analysis of serotype K1 Pseudomonas aeruginosa strains isolated from liver abscess patients and noninfectious stool samples. J. Clin. Microbiol. 49(11), 3761–3765. Stover, C.K., Pham, X.Q., Erwin, A.L., Mizoguchi, S.D., Warrener, P., Hickey, M.J. and Olson, M.V. 2000. Complete genome sequence of Pseudomonas aeruginosa PAO1. Nature 406, 959–964. | ||
| How to Cite this Article |
| Pubmed Style Alshammari MMM, Kamil SA, Gaber HH, Faja OM, Al-zuhairi ZA, Muhana BMMA. Determinants of genetic diversity, multidrug resistance, and virulence of Pseudomonas aeruginosa isolated from aquaculture fish in Central Iraq. Open Vet. J.. 2026; 16(5): 2766-2780. doi:10.5455/OVJ.2026.v16.i5.19 Web Style Alshammari MMM, Kamil SA, Gaber HH, Faja OM, Al-zuhairi ZA, Muhana BMMA. Determinants of genetic diversity, multidrug resistance, and virulence of Pseudomonas aeruginosa isolated from aquaculture fish in Central Iraq. https://www.openveterinaryjournal.com/?mno=300083 [Access: June 26, 2026]. doi:10.5455/OVJ.2026.v16.i5.19 AMA (American Medical Association) Style Alshammari MMM, Kamil SA, Gaber HH, Faja OM, Al-zuhairi ZA, Muhana BMMA. Determinants of genetic diversity, multidrug resistance, and virulence of Pseudomonas aeruginosa isolated from aquaculture fish in Central Iraq. Open Vet. J.. 2026; 16(5): 2766-2780. doi:10.5455/OVJ.2026.v16.i5.19 Vancouver/ICMJE Style Alshammari MMM, Kamil SA, Gaber HH, Faja OM, Al-zuhairi ZA, Muhana BMMA. Determinants of genetic diversity, multidrug resistance, and virulence of Pseudomonas aeruginosa isolated from aquaculture fish in Central Iraq. Open Vet. J.. (2026), [cited June 26, 2026]; 16(5): 2766-2780. doi:10.5455/OVJ.2026.v16.i5.19 Harvard Style Alshammari, M. M. M., Kamil, . S. A., Gaber, . H. H., Faja, . O. M., Al-zuhairi, . Z. A. & Muhana, . B. M. M. A. (2026) Determinants of genetic diversity, multidrug resistance, and virulence of Pseudomonas aeruginosa isolated from aquaculture fish in Central Iraq. Open Vet. J., 16 (5), 2766-2780. doi:10.5455/OVJ.2026.v16.i5.19 Turabian Style Alshammari, Majida Malik Meteab, Shahad Ali Kamil, Huda Hassan Gaber, Orooba Meteab Faja, Zahira A. Al-zuhairi, and Balsam Miri Mizher Al Muhana. 2026. Determinants of genetic diversity, multidrug resistance, and virulence of Pseudomonas aeruginosa isolated from aquaculture fish in Central Iraq. Open Veterinary Journal, 16 (5), 2766-2780. doi:10.5455/OVJ.2026.v16.i5.19 Chicago Style Alshammari, Majida Malik Meteab, Shahad Ali Kamil, Huda Hassan Gaber, Orooba Meteab Faja, Zahira A. Al-zuhairi, and Balsam Miri Mizher Al Muhana. "Determinants of genetic diversity, multidrug resistance, and virulence of Pseudomonas aeruginosa isolated from aquaculture fish in Central Iraq." Open Veterinary Journal 16 (2026), 2766-2780. doi:10.5455/OVJ.2026.v16.i5.19 MLA (The Modern Language Association) Style Alshammari, Majida Malik Meteab, Shahad Ali Kamil, Huda Hassan Gaber, Orooba Meteab Faja, Zahira A. Al-zuhairi, and Balsam Miri Mizher Al Muhana. "Determinants of genetic diversity, multidrug resistance, and virulence of Pseudomonas aeruginosa isolated from aquaculture fish in Central Iraq." Open Veterinary Journal 16.5 (2026), 2766-2780. Print. doi:10.5455/OVJ.2026.v16.i5.19 APA (American Psychological Association) Style Alshammari, M. M. M., Kamil, . S. A., Gaber, . H. H., Faja, . O. M., Al-zuhairi, . Z. A. & Muhana, . B. M. M. A. (2026) Determinants of genetic diversity, multidrug resistance, and virulence of Pseudomonas aeruginosa isolated from aquaculture fish in Central Iraq. Open Veterinary Journal, 16 (5), 2766-2780. doi:10.5455/OVJ.2026.v16.i5.19 |

