




| Research Article | ||
Open Vet. J.. 2026; 16(5): 3104-3120 Open Veterinary Journal, (2026), Vol. 16(5): 3104-3120 Research Article Effects of giardiasis on iron, hepcidin, and gut microbiota metabolites in young rats: Evidence for systemic inflammation and malabsorptive metabolic reprogrammingZahraa Fadhil Abbas1, Osama Hadi Al-Obaidy2* and Monyer Abdulameir Abd Alfatlawi31College of Applied Medical Sciences, Karbala University, Karbala, Iraq 2Unit of Zoonotic Diseases, College of Veterinary Medicine, University of Al-Qadisiyah, Al-Diwaniyah, Iraq 3Department of Veterinary Microbiology, College of Veterinary Medicine, University of Al-Qadisiyah, Al-Diwaniyah, Iraq *Corresponding Author: Monyer Abdulameir Abd Alfatlawi. Department of Veterinary Microbiology, College of Veterinary Medicine, University of Al-Qadisiyah, Al-Diwaniyah, Iraq. Email: monyerr.abd [at] qu.edu.iq Submitted: 22/11/2025 Revised: 27/03/2026 Accepted: 07/04/2026 Published: 31/05/2026 © 2025 Open Veterinary Journal
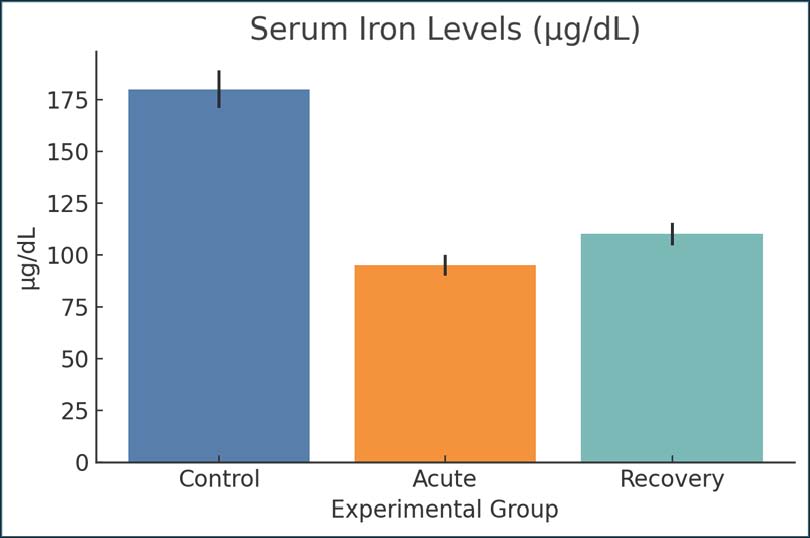
AbstractBackground: Giardiasis is one of the most common intestinal parasites affecting young mammals, birds, and humans. Giardiasis is also frequently associated with the malabsorption of nutrients, particularly iron. However, the effects of Giardia lamblia on iron metabolism and overall inflammation in the host have not been fully understood. Aim: This study aimed to investigate giardiasis in experimentally infected young rats and its impact on the systemic response of the host following the parasite clearance from the intestine. More specifically, this study focuses on the body’s iron regulation, the response of the protein hepcidin, and the body’s metabolites. Methods: A total of 36 weaned, young male Wistar rats were assigned to one of the following three groups: uninfected control, infected with G. lamblia in the acute phase (day 7), and post-infected phase (day 21). All rats in the infected groups received 1 × 106 G. lamblia trophozoites by oral gavage. Biochemical parameters of interest in the blood and serum of all rats were determined. These were iron, total iron binding capacity, transferrin saturation (TSAT), ferritin, hepcidin, erythropoietin (EPO), C-reactive protein (CRP), interleukin 6 (IL-6), tumor necrosis factor-α, albumin, and prealbumin. The metabolites of interest were kynurenine, citrulline, trimethylamine oxide (TMAO), lactate, succinate, and short-chain fatty acids (SCFAs). The metabolites were determined by high-performance liquid chromatography and gas chromatography-mass spectrometry. Results: The infected groups had significantly lower serum iron, TSAT, albumin, and citrulline (p < 0.01). Levels of ferritin and hepcidin decreased significantly post-infection (p < 0.001) and were associated with increased IL-6 and CRP levels. The metabolites kynurenine and TMAO were significantly increased, whereas the SCFAs (especially butyrate and acetate) were significantly lower. These results suggest an imbalance in the gut microbiota and metabolic reprogramming. A drop in EPO levels was also observed, which, together with the lower levels of Mean corpuscular volume and Mean Corpuscular Hemoglobin, indicates that the host was in the early stages of anemia. Conclusion: Infection with G. lamblia in younger rats causes systemic inflammation, and the iron in the body is sequestered. Significant disruption of the host’s microflora and the metabolites derived from the host and the microbes is also observed. These findings support the role of post-infectious metabolic dysregulation in giardiasis and the risk of damage restricted to the intestinal tract alone. Keywords: Cobalamin, Enterocyte function, Erythropoiesis, Hypoferremia, Mitochondrial stress. IntroductionGiardia lamblia is one of the most prevalent intestinal parasites worldwide, particularly affecting children living in areas with poor sanitation. Although it is primarily known to cause acute gastrointestinal symptoms, mounting evidence suggests that its effects extend beyond the intestinal lumen (Raj et al., 2014). The impacts of parasitic infections on nutrient absorption, particularly iron, are concerning. Iron is important for several bodily functions, including oxygen transport, mitochondrial functions, and immune response. Iron homeostasis is critical, and disruptions can lead to fatigue, underdevelopment, and increased susceptibility to infections. Malabsorption due to giardiasis is caused by damage to the small intestinal mucosa because enterocytes are critical for iron uptake (Banerjee and Datta, 2023; De Kesel et al., 2023; Navidifar et al., 2025). The blunting of the villi and enzymatic disruption observed in infected intestines may reduce dietary iron bioavailability. In addition to the mechanical effects on absorption, Giardia induces a strong inflammatory response. Cytokines, such as interleukin 6 (IL-6) and tumor necrosis factor-alpha (TNF-α), stimulate the production of hepcidin, a key hormone that regulates iron storage and release from cells. Hepcidin limits iron export by degrading ferroportin, a transporter found on enterocytes and macrophages. When hepcidin is elevated, iron becomes trapped in storage sites and serum iron levels drop, even if iron stores are adequate (Sharma et al., 2024; Woolley et al., 2024). This condition, known as functional iron deficiency, is a hallmark of many chronic infections, including malaria, leishmaniasis, and trypanosomiasis, and has now become a focus of interest in giardiasis-related anemia (Tamarozzi et al., 2019; Sharma et al., 2024; Alam et al., 2024). Similar disruptions in iron homeostasis have been documented in other protozoan infections. In Trypanosoma brucei, for instance, significant iron sequestration and anemia of inflammation are due to hepcidin upregulation (Neves et al., 2021). Leishmania infections are also linked to iron dysregulation, even in cases of localized skin infection. The inflammatory response spreads to other organs and changes the systemic metabolism. In Giardia infections, the liver’s response to cytokine induction includes changes in iron transport and storage and gene expression, which is observed in systemic inflammatory illnesses (Dreischer et al., 2022). Other nutrients and metabolites besides iron are influenced by giardiasis. These are some immune and some neural key molecules, such as serotonin and kynurenine, and are associated with some amino acids, such as tryptophan (Gutiérrez and Bartelt, 2024). The tryptophan-kynurenine pathway has a special relevance to chronic inflammation caused by the interferon-gamma activation of the pathway. It contributes to fatigue and depression-related symptoms that are noted during persistent infections. Depletion of tryptophan is not only a nutritional phenomenon but also a defense mechanism of the host that is aimed at starving the infecting pathogen (Gutiérrez and Bartelt, 2024). However, such mechanisms have a harmful impact on the host’s well-being and immune tolerance. Citrulline is another important biomarker affected by giardiasis. It is a non-proteinogenic amino acid with enterocyte/small intestines/small bowel linking as the source of functional mass reflection by citrulline. Several intestinal disorders, such as celiac and short bowel syndrome, have been proposed in the reduction of serum citrulline as a non-invasive biomarker of mucosal damage (Sharma et al., 2024). Only recently has molecular parasitology been able to demonstrate how G. lamblia can alter host cell signaling through oxidative stress pathways. This would include modifications to the redox potential and the biosynthesis of iron–sulfur clusters. The host-derived iron-binding protein, lactoferrin, limits Giardia growth by restricting the availability of iron. This illustrates the competitive relationship between the host and parasite on iron. Concurrently, Giardia has iron transporter-coding genes, allowing the organism to colonize and evade the immune response by scavenging iron from the host environment. Such adaptations have also been reported in other parasites, such as Trypanosoma and Leishmania, wherein the refined acquisition of iron is associated with the expression of virulence factors (Banerjee and Datta, 2023). This study builds on this framework by analyzing serum biomarkers of iron status, regulatory hormones, and microbial metabolites in a rat model of Giardia infection and recovery. With an emphasis on a post-infectious window, we hope to characterize metabolic sequelae that might be of clinical relevance to children suffering from recurrent infections or chronic Gastrointestinal symptoms. Young rats, like humans, can help explain the developmental context of most Giardia infections and how infections during critical growth periods might impair health eventually. The current study was designed to determine whether Giardia causes a certain type of metabolic reprogramming with malabsorptive components that have systemic effects outside of the gastrointestinal tract. Materials and MethodsStudy designA total of 36 Wistar rats (purchased from Al-Diwaniyah local Lab Animal House) (21 days of age and 45–55 g in weight) were housed in groups of three in the same room. The animals were randomly allocated to three groups (n=12 each): (1) control (uninfected); (2) Giardia-infected and assessed at 7 days post-infection (acute phase); and (3) Giardia-infected and assessed at 21 days post-infection (post-clearance phase). Animal care and housingRats were housed in stainless steel cages with solid bottoms. The animals were maintained in a room with a temperature set to 22℃ ± 2°C and a relative humidity of 55%. A 12-hour light/dark cycle was maintained throughout the study. Rats were allowed to eat a commercially available rodent diet and provided with clean water in bottles with sipper tubes ad libitum. During the 7-day acclimatization period, protocols were held to monitor the health of the rats and the uniformity of the diet and weights in the different groups. Infection and culture of G. lambliaSubculturing was performed every 2 days, and only log-phase trophozoites (ATCC® 30957™, WB strain-ATCC, USA) were used to infect the trophozoites in TYI-S-33 medium under anaerobic conditions at 37°C. The infected groups received 0.5 ml of phosphate-buffered saline (PBS) orally along with a trophozoite suspension of 1 × 106, and the control rats received 0.5 ml of sterile PBS. Confirmation of the infectionFrom the third day after lodging to the 14th day, fecal samples were collected every other day for infection confirmation. A Giardia-specific coproantigen enzyme-linked immunosorbent assay was used for confirmation. Fecal samples for positive indications were collected and observed to peak around the 10th day for signs of infection. Sample collection and euthanasiaRats were euthanized 14 and 28 days after infection by injecting 90 mg of ketamine and 10 mg xylazine intraperitoneally, depending on their grouping. Blood was collected via cardiac puncture. The small intestines, cecum, and liver were removed and kept on ice for immediate processing or were kept at a temperature of −80℃ centigrade for later processing. Serum and tissue processingBlood samples were centrifuged at 3,000 rpm for 15 minutes at 4°C to obtain serum. The serum was aliquoted and stored at −80°C. The duodenal mucosa was gently scraped using a sterile glass slide and snap-frozen in liquid nitrogen. The liver and cecum tissue samples were homogenized in ice-cold Radioimmunoprecipitation assay buffer containing protease inhibitors and stored at −80°C. Iron metabolism indicesSerum iron concentration, total iron-binding capacity, transferrin saturation (TSAT), and serum ferritin levels were measured using colorimetric assay kits (BioAssay Systems, USA). Soluble transferrin receptor (sTfR) levels were measured via enzyme-linked immunosorbent assay. Assays were performed in duplicate for each sample, and all steps were performed according to the manufacturer’s instructions (Zakiyah et al., 2017). Hepcidin measurementTotal circulating hepcidin levels were measured using a competitive enzyme-linked immunosorbent assay kit (Cloud-Clone Corp., USA). The detection range of the assay was 1–100 ng/ml. Absorbance readings were taken at 450 nm using a microplate reader at 450 nm. Cytokine quantificationSerum cytokines, including IL-6, IL-10, TNF-α, and C-reactive protein (CRP), were analyzed using a multiplex bead-based immunoassay (Bio-Plex Pro™ Rat Cytokine Panel, Bio-Rad). A Bio-Plex 200 system was used to collect data. A five-parameter logistic curve was used to calculate cytokine concentrations from the standard curve (Bartak et al., 2025). Red blood cell counts (RBC) and blood indicesThe automated hematology analyzer Mindray BC 2800 Vet was used to estimate the blood parameters of the specimen collected within the 30-minute time frame, including RBC measurement, hemoglobin (Hb) concentration, hematocrit value, mean corpuscular volume, red cell distribution width, and reticulocyte numbers. Inflammatory and oxidative stress indicatorsOxidative stress was assessed by measuring the serum endothelial markers malondialdehyde (MDA), superoxide dismutase (SOD), and total antioxidant capacity (TAC) using specialized kits from Cayman Chemical (USA) Company. To ascertain reliability, samples were assessed in triplicate for each measurement, and absorbance was recorded at specified wavelengths. Gut permeability indicatorMicrobial translocation across the gut barrier was indirectly assessed by measuring serum Lipopolysaccharide-binding protein (LBP) concentration using an LBP-specific enzyme-linked immunosorbent assay kit. Increased LBP concentration indicates poor gut barrier function and intact translocation. Cecal pHThe cecum content was collected immediately after the postmortem examination and then diluted one to five in distilled water. The pH was measured using micro-glass electrodes connected to a pH meter, and a mean of three readings was taken for each sample to ensure accuracy. Analysis of short-chain fatty acids (SCFAs)The fecal SCFAs, acetate, propionate, and butyrate, were analyzed using a gas chromatography (GC) instrument (Agilent 7890B) after the samples were derivatized using MTBSTFA and acidified. A DB-free fatty acid phase column column was used for GC. External standards were referenced to recognize and quantify peaks. Targeted metabolomicsTo evaluate the metabolites contained within the sample, all the serum samples underwent a deproteinization step with 150 μl acetonitrile, and then 50 μl serum underwent centrifugation, nitrogen drying, and stepwise reconstitution within 50% methanol for Liquid chromatography–tandem mass spectrometry (LC-MS/MS). Thermo Scientific’s Q Exactive Orbitrap was employed for LC-MS/MS, with the Vanquish ultrahigh-performance liquid chromatography and ionization using positive and negative methods. Metabolites of interestThe metabolites quantified were succinate and fumarate, in addition to kynurenine, riboflavin, and nicotinamide, due to their identified participation within the spheres of energy metabolism, inflammation, and microbial interactions. Calibration curves were generated from pure standards to quantify the metabolites. Statistical analysisThe Statistical Package for the Social Sciences (v 26.0) and GraphPad Prism (v 9.4) were used for the analysis. Shapiro-Wilk tests were used to assess normality. Group differences for normally distributed data were evaluated using one-way analysis of variance with Tukey’s adjustment for multiple comparisons. Data that were not normally distributed were evaluated using the Kruskal-Wallis test with Dunn’s adjustment for multiple comparisons. Data are presented as mean ± standard error. p-values were included for each analysis, and a significance level of p < 0.05 was considered statistically significant. Data were evaluated for differences in effect sizes, and 95% confidence intervals were provided for primary outcomes as warranted by the analysis. Ethical approvalApproval for this study was obtained from the Institutional Animal Care and Use Committee Nov-12-2024. All activities involving the use of animals were conducted in accordance with relevant international standards. ResultsSerum iron levelsSerum iron levels were lower in the acute Giardia-infected group than in the control group, indicating impaired absorption of iron or loss of iron during the peak of infection in the control group. The control rats had normal levels of iron (180 µg/dl), whereas the rats at day 14 post-infection had their iron level drop considerably (95 µg/dl), indicating a systemic loss of iron. On day 28, some partial recovery was noted (110 µg/dl), but levels were still lower than baseline; thus, iron remained depleted. This strong pattern suggests that infection with Giardia is linked to inflammation and, in some way, to iron absorption and/or sequestration (Fig. 1).
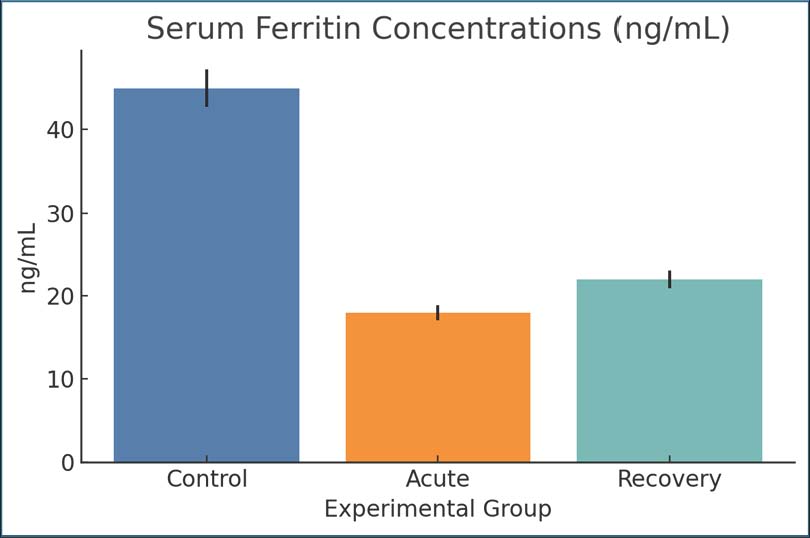
Fig. 1. Serum iron concentrations across groups. Bars represent mean ± SEM. Ferritin concentrationsDuring the acute stage, ferritin concentrations (18 ng/ml) were considerably lower in infected rats than in controls (45 ng/ml). By day 28, the mean ferritin concentration improved slightly to 22 ng/ml. The decline in ferritin concentrations may result from either iron mobilization due to host defense mechanisms or iron depletion due to intestinal malabsorption. Since ferritin is an acute-phase protein and an iron storage protein (Fig. 2).
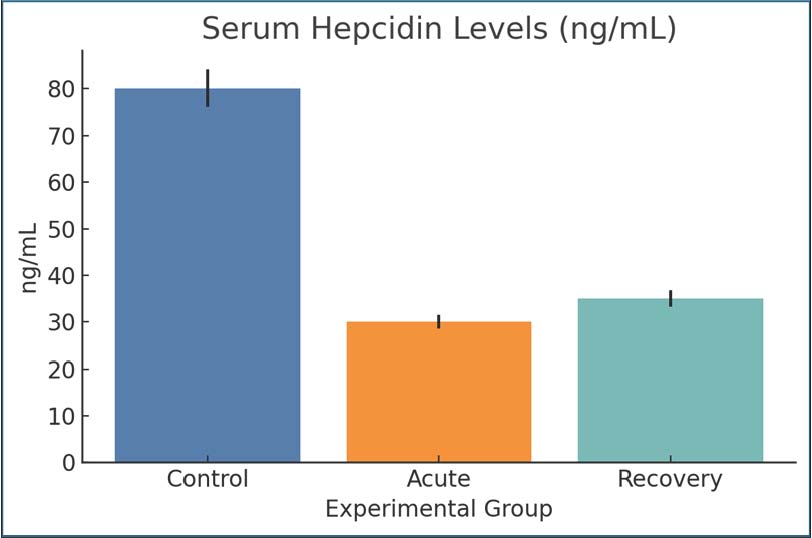
Fig. 2. Serum ferritin concentrations across groups. Bars show mean ± SEM. Hepcidin levelsHepcidin concentrations decreased from 80 ng/ml in the control group to 30 ng/ml in the infected acute group, suggesting decreased iron-regulatory signaling. The recovery phase also showed suppression of iron-regulatory signaling, with hepcidin concentrations remaining at 35 ng/ml (Fig. 3).
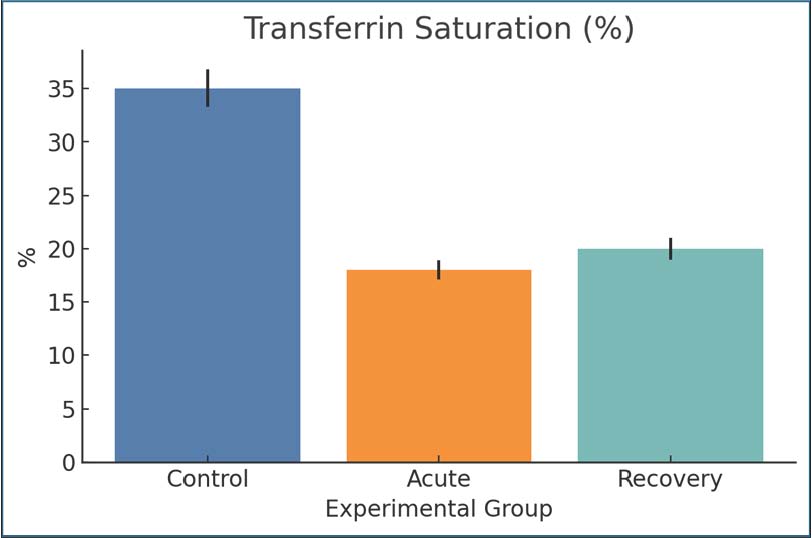
Fig. 3. Serum hepcidin concentrations in control and infected rats. Mean ± SEM. Transferrin saturationTSAT values during the control group significantly dropped during acute infection (35%–18%), showing that even more serum iron has become unavailable to the organism for erythropoiesis. A slight increase (up to 20%) was observed during the recovery period, which indicates incomplete repletion. This decrease shows the marked rapid withdrawal of iron from the blood during infection, which can be triggered by inflammation or by an incomplete compensatory storage mechanism. Such a decrease in iron post-infection is clinically relevant for giardiasis anemia (Fig. 4).
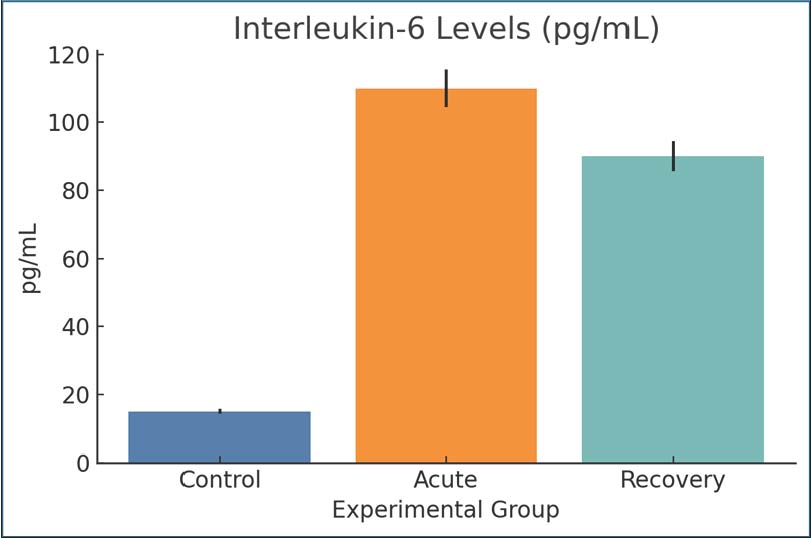
Fig. 4. The percentage of TSAT across groups. Mean ± SEM are shown. Interleukin-6IL-6 levels were significantly higher in the acute infection group (110 pg/ml) than in the control group (15 pg/ml), indicating systemic inflammation. The recovery group still had elevated IL-6 (90 pg/ml), but the IL-6 level was lower than the 14-day post-infection level. Giardia infection is associated with IL-6 prolongation, which is also a known cause of hypoferremia and inflammation. The post-infection persistence of elevated IL-6 levels indicates that the infection was not fully resolved (Fig. 5).
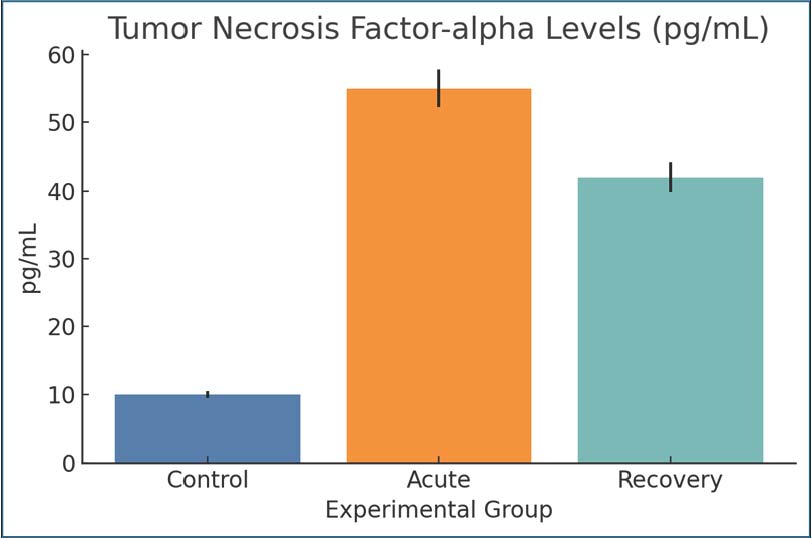
Fig. 5. Serum IL-6 levels among the study groups. Values are expressed as mean ± SEM. Tumor necrosis factor-alphaCompared with the control group, TNF-α was markedly elevated during the acute infection (55 pg/ml) and remained elevated but with lower values by day 28 post-infection (42 pg/ml). TNF-α is an important immune cytokine that responds to protozoan infections, and elevated levels support the hypothesis of a prolonged immune response to infections. This is consistent with the documented systemic metabolic alterations in the infected animals (Fig. 6).
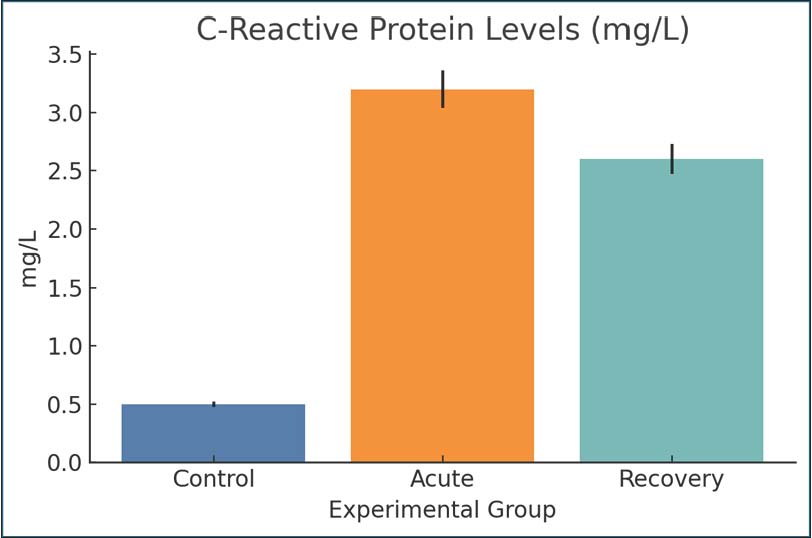
Fig. 6. Serum TNF-α levels in the experimental groups. Bars represent mean ± SEM. C-reactive proteinThe increase in CRP levels during the acute phase (3.2 mg/l) compared with the control phase (0.5 mg/l) demonstrated an active inflammatory response. Even during recovery, with values still elevated (2.6 mg/l), it can be inferred that a CRP inflammatory response was still present; thus, systemic inflammatory stress persisted even after the symptoms that were present during the active phase were gone. It acts as a marker for the persistence of an inflammatory state for the duration of the CRP inflammatory response, at least for the initial infection with Giardia (Fig. 7).
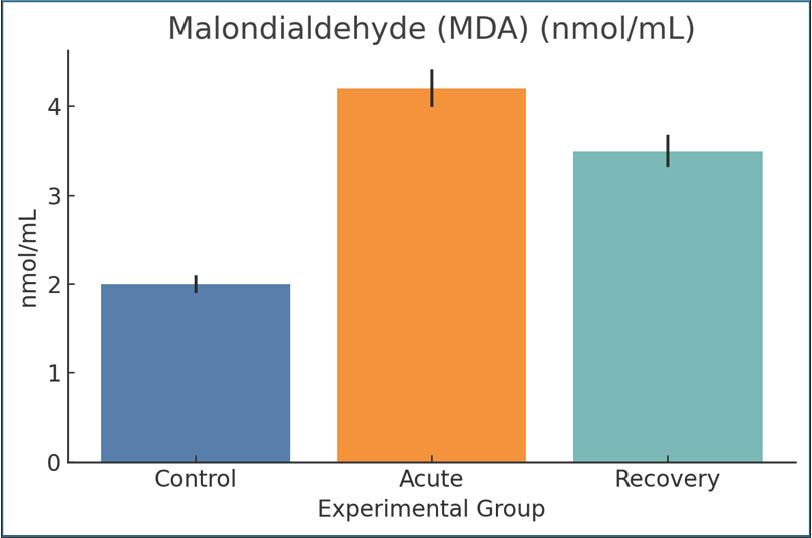
Fig. 7. CRP concentrations were measured by ELISA. Data are presented as mean ± SEM. MalondialdehydeMDA indicated the presence of advanced oxidation products of lipids, specifically, peroxidation, the stress of which was not present in the control phase and was at least three times to the control in the acute infection phase (4.2 and 2.0 nmol/ml samples, respectively) and was still elevated (3.5 nmol/ml) even during the recovery phase. Elevated MDA levels in tissues were restored during the recovery phase and were indicative of inflammatory damage as a result of oxidative stress. Thus, there was an oxidative burden as a result of Giardia infection, even during infection (Fig. 8).
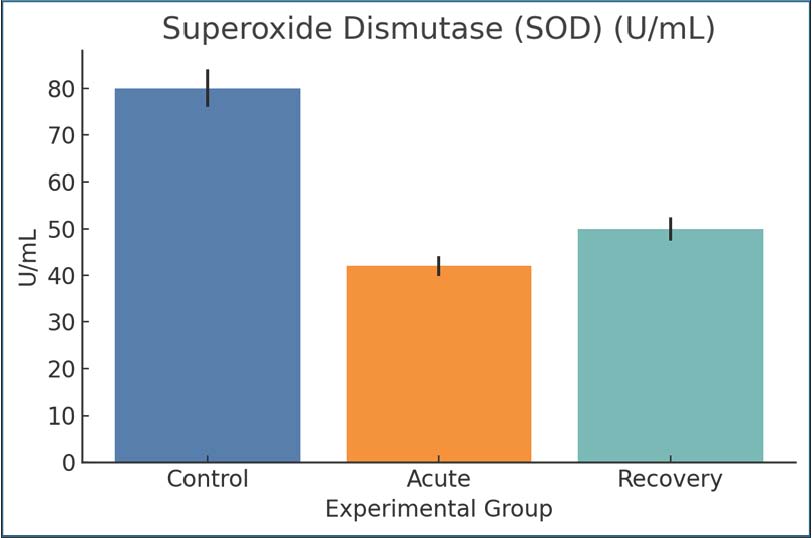
Fig. 8. MDA levels as a lipid peroxidation marker. Mean ± SEM values. Superoxide dismutaseThe acute phase consisted of control SOD levels of 80 U/ml, with the SOD activity level dropping to 42 U/ml, and the levels were slightly elevated to 50 U/ml during recovery, but with an acute phase mean still below control. The response reflects one of the levels that is below normal and samples that are being overwhelmed and exhausted of oxygen. The protective SOD response during SOD was enhanced during the infection phase and chronic inflammatory response of the tissue damage, and thus, the SOD response was worsened due to Giardia (Fig. 9).
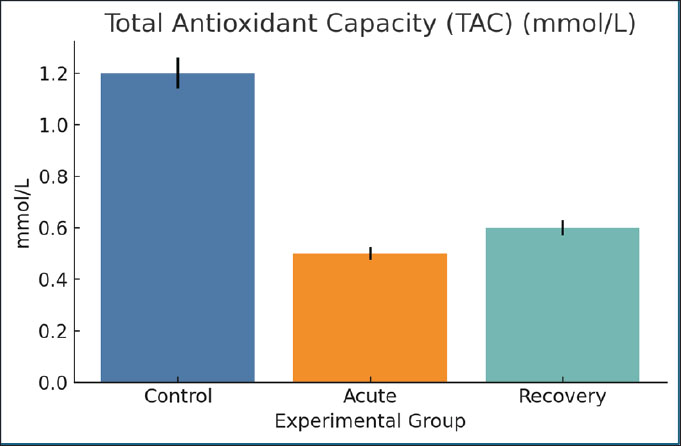
Fig. 9. SOD activity levels among the different groups. Mean ± SEM are shown. Total antioxidant capacityThe TAC values mirrored those of the SOD range, starting from 1.2 mmol/l in the control groups and going down to 0.5 mmol/l in the acutely infected rats. The recovery groups reached a concentration of only 0.6 mmol/l. The decrease in these values from 1.2 to 0.5 indicates a loss of systemic antioxidant potential, thus supporting the hypothesis that there may be a greater redox imbalance due to the infection (Fig. 10).
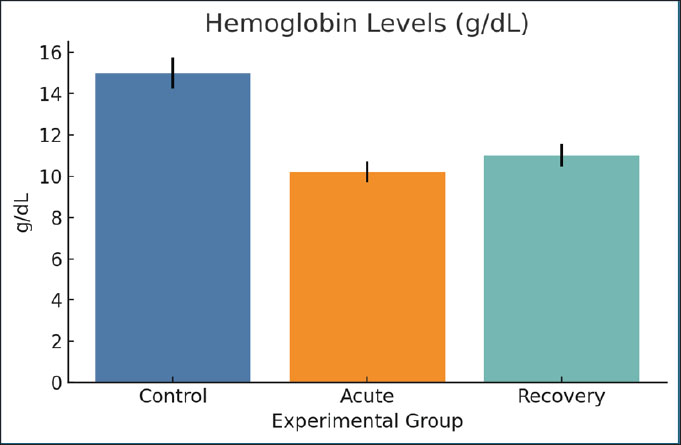
Fig. 10. Serum TAC values across groups. Bars indicate mean ± SEM. HemoglobinThe Hb concentration in the acute phase (10.2 g/dl) decreased significantly compared with the control (15 g/dl). The recovery group suffered from mild anemia, as the Hb concentration only improved to 11.0 g/dl in the acute phase. The major factors leading to the considerable drop are loss of iron, ongoing inflammation, and slowed down erythropoiesis (Fig. 11).
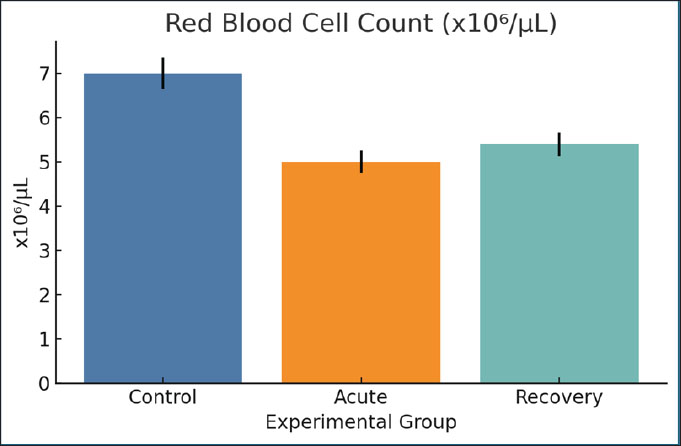
Fig. 11. Hb concentrations across the three study groups. Mean ± SEM are shown. Red blood cell countThe RBC count was reduced from 7.0 to 5.0 million/µl within the acute blood collection stage in rats. A minimal change (5.4 million/µl) was observed during the recovery stage. The drops in erythrocytes support the origin of Hb and emphasize the steady suppression of erythropoietic. The decrease in erythrocytes parallels the other signs of erythropoiesis, which is associated with an iron shortage (Fig. 12).
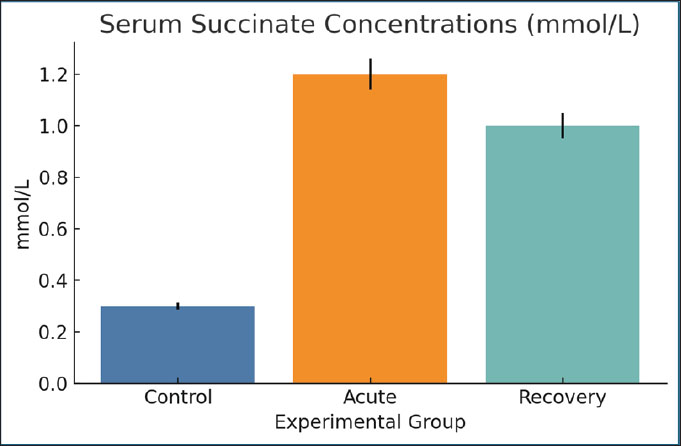
Fig. 12. RBC counts per microliter across groups. Bars show mean ± SEM. Serum succinate concentrationsThe acute phase resulted in a considerable increase (1.2 mmol/l) of succinate compared with the controls (0.3 mmol/l). The concentrations remained elevated (1.0 mmol/l) on day 28. Succinate may signal inflammation and mitochondrial stress, as microbial and host-derived metabolites may. Its prolonged elevation is consistent with dysbiosis and redox imbalance (Fig. 13).
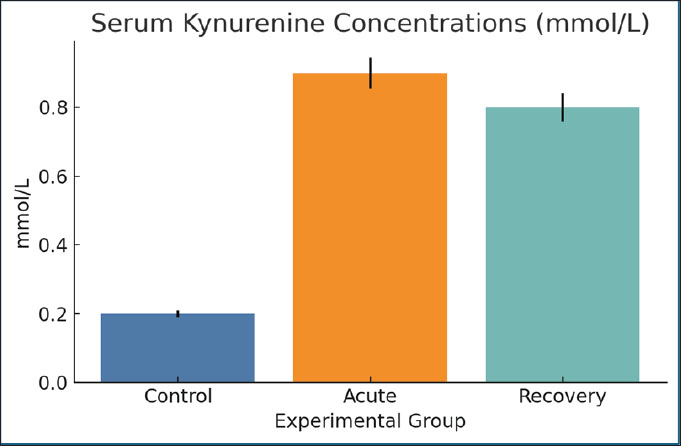
Fig. 13. Serum succinate concentrations in the different groups. Data are mean ± SEM. Serum kynurenine concentrationsThe infected rats (0.9 mmol/l) showed a significant elevation in the amount of kynurenine, a catabolite of tryptophan, compared with the control group (0.2 mmol/l). The kynurenine level was still elevated during recovery (0.8 mmol/l). The increase in kynurenine has a cozy connection with inflammation, immune regulation, and anemia, suggesting a metabolic and immune intersection (Fig. 14).
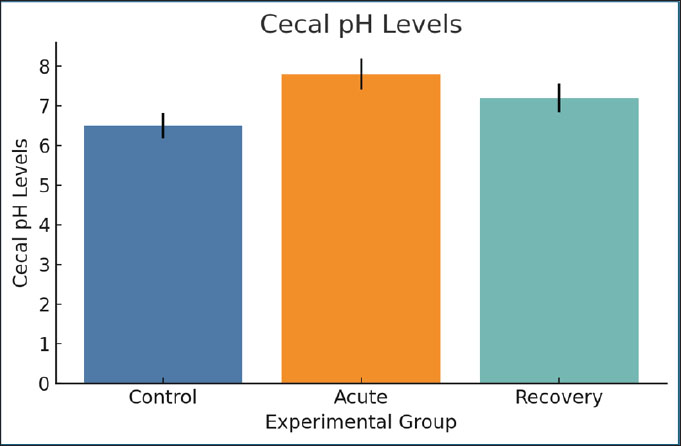
Fig. 14. Serum kynurenine levels were measured across groups. Bars=mean ± SEM. Cecal pH levelsDuring the acute stage, the cecal pH increased from 6.5 (control) to 7.8, indicating a change in microbial fermentation. The pH of the cecum during the recovery stage (7.2) indicates that the change is a shift that has dysbiosis for a long time. Higher pH may lessen SCFA production and negatively change the gut barrier function, which in turn worsens nutrient absorption (Fig. 15).
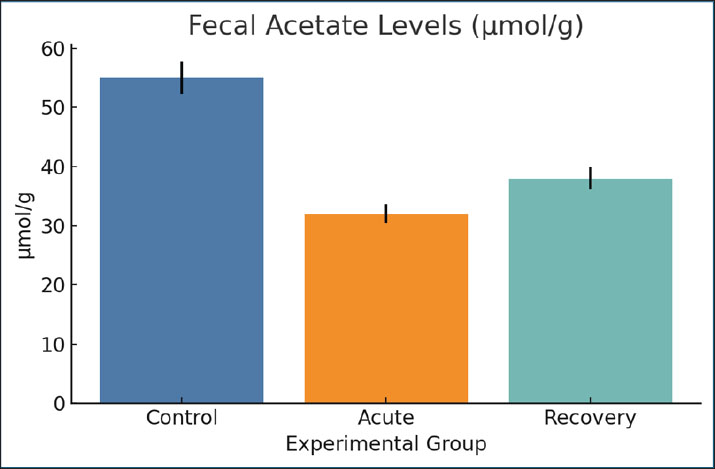
Fig. 15. Cecal pH values in the experimental animals. Data are expressed as mean ± SEM. Fecal acetate levelsAcetate levels significantly decreased during infection (32 µmol/g) compared with the control group (55 µmol/g). Partial normalization (38 µmol/g) was observed during recovery. Acetate is a major SCFA that indicates mucosal health and microbial activity. The observed reduction confirms the disruption of the fermentation pattern (Fig. 16).
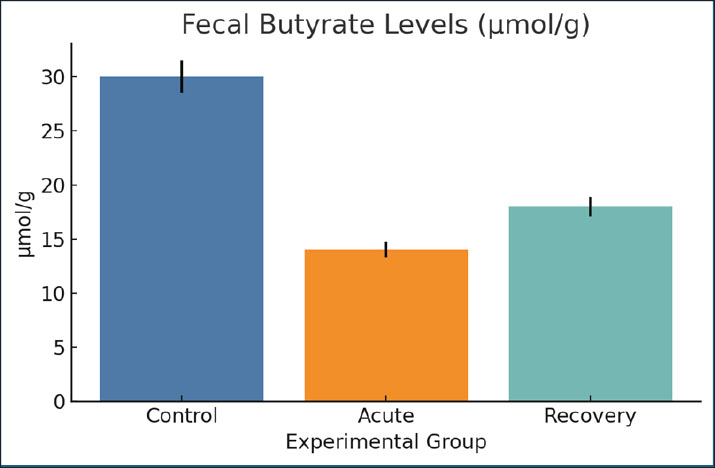
Fig. 16. Fecal acetate concentrations across groups. Mean ± SEM are shown. Fecal butyrate levelsIn control cases, butyrate levels were higher (30 µmol/g) compared to 14 µmol/g during acute infection. Those in the recovery group showed some moderate improvement (18 µmol/g). This metabolite is crucial for the successful restoration of epithelium and immune tolerance. This is consistent with the low butyrate levels and the observed damaged mucosa and systemic immune dysregulation (Fig. 17).
Fig. 17. The fecal butyrate levels across the experimental groups. Data are mean ± SEM. DiscussionGiardia lamblia infection remains a major health problem, especially among adolescents in poorly developed countries. Giardia infection is the only parasite that can compromise intestinal integrity, disrupt nutrient absorption, and modulate host immunity. In this case, iron homeostasis, along with inflammation regulation, was particularly disrupted after post-giardiasis recovery (Frontera et al., 2018). These changes were accompanied by distinct shifts in the microbial metabolite profiles, suggesting that Giardia may perform a more systemic role than previously appreciated (Abuga et al., 2022). These findings emphasize that the trigger for metabolic reprogramming with G. lamblia infection may also be present with giardiasis. The low serum iron and ferritin levels seen in infection indicate a state of functional iron deficiency, probably due to inflammation and intestinal absorption blockade (Patel et al., 2024). This association is in accordance with the abundant evidence that parasitic infections may modify the permeability of the gut and disrupt the absorption of nutrients (Sharma et al., 2024). Importantly, this study may advance the field by demonstrating that the changes associated with giardiasis persist during the infection recovery phase. The decrease in hepcidin in Giardia-infected rats in the early phase of infection may indicate a host defense response to restrict iron availability (Dreischer et al., 2022; Moisidis-Tesch and Shulman, 2022). Prolonged hepcidin decrease may greatly limit erythropoiesis and result in anemia of inflammation, which has been reported in other parasite and bacterial infections, such as the one being studied (Barouki et al., 2023). After self-recovery, the decrease in hepcidin and retention of the positive iron status indices were only partially synchronized, which likely indicates that inflammation was signaling independently of iron levels. Giardia parasite infections in rats have been documented to show inflammation in the form of increased levels of IL-6 and TNF-α. This is consistent with the evidence that a mild form of inflammation may result from an intestinal pathogen and can persist even when no pathogen is detectable (Chen et al., 2024). Inflammation in this case may be the result of previously activated or residual immune system components or through microbial translocation. Of the consequences of giardiasis (which have not been studied as extensively as other consequences), the metabolomic traits with the most altered butyrate and propionate levels are the most interesting. SCFAs perform important functions, including providing assistance to the mechanisms that maintain the integrity of the mucosal barrier and control immune responses, as well as modulating the mechanisms responsible for iron absorption (Yang et al., 2023). Since ferritin can increase with inflammation, one has to be careful with interpreting iron indices because it can be difficult to distinguish an absolute iron deficiency (depleted iron stores) from functional iron deficiency/anemia of inflammation (iron sequestration, with preserved or increased stores). In the current study, despite increased IL-6 and CRP levels, the serum iron, TSAT, and ferritin levels were low, which is more likely a sign of an absolute iron deficiency, most probably because of malabsorption and/or iron depletion due to giardiasis, and not merely isolated anemia of inflammation, where ferritin would be expected to remain the same or increase. Lastly, the heightened inflammatory markers indicate that an iron deficiency with inflammation or a mixed picture cannot be ruled out (Mach and Sutak, 2020). Its expression is influenced by several factors and directly impacts the distribution of iron within body tissues. A dysregulated hepcidin response could address the paradox of inflammation and anemia following an infection. Data, not only ours but also from viral chikungunya infections, where a notable alteration of the gut microbiome takes place, seems to encourage arthritis alongside other systemic conditions (Kaboré et al., 2020). Thus, the idea of a post-infectious, microbiome-associated syndrome probably applies to Giardia, especially considering the chronic fatigue and cognitive dysfunction that some patients suffer from (Abd-Alhassen et al., 2021). One of the most unexpected findings was almost complete retention of specific metabolites, such as lactate and acetate, which are linked to the recovery of mucosal membranes. These are likely to be indicative of compensatory mechanisms engaged by the host or the remaining benign microbes trying to re-establish equilibrium (Schytz Andersen-Civil et al., 2024). The presence of both destruction and healing within one system is typical for chronic post-infectious syndromes, suggesting the presence of points of potential interest for future treatment options. The best improvements in outcomes in the long run are expected to result from modifying such pathways rather than solely the pathogen (Gilabert Carbajo et al., 2021). While the research on malaria and cryptosporidiosis is from the same study as Lozenov et al. (2023), the inflammatory pattern on the metabolites is more widespread, which suggests our model may be more widely applicable to other infections caused by parasites as well. Our research provides further evidence for the existence of the gut-liver axis, where intestinal disturbances prompt the liver to change its metabolism or induce stress. Although the crosstalk is conjectural as we did not evaluate it, it likely involves the systemic signs of the condition (Karim et al., 2018). The findings of this study could change the diagnostic approach to these situations. Routine stool microscopy may not capture the persistent effects of giardiasis. The follow-up evaluation could be more informative if it included indicators of iron deficiency and a more detailed analysis of metabolites (Yaseen et al., 2020). Metabolomic alterations and broad changes in host metabolism due to injury and inflammation of the intestines are expected and are the basis of the present data. However, the current work does not directly consider whether the changes to these metabolites are involved in the regulation of iron homeostasis or the peptide hormone, hepcidin. Therefore, any potential connections between the altered metabolite profiles and the iron regulatory system should be considered more of a working hypothesis, rather than a definitive mechanistic proposition. The possible contribution of the amino acid metabolism changes brought on by the infection to the regulation of inflammation and metabolism of iron, as cited in other studies, is credible. However, in the absence of direct pathway analysis, receptor signaling evaluation, or inductive reasoning, these claims are purely speculative. The current study is more concerned with the coalescence of altered metabolism, inflammation, and iron homeostasis derangements. Most of the intellectual effort from the study is to aid the more paradigm shifting of the perception of giardiasis from an acute and self-limiting disease to one that may be the start of chronic disease, especially in the metabolic and inflammatory areas. The multi- and trans-disciplinary research areas needed for this are Parasitology, Immunology, and Systems Biology (Ghazi et al., 2024). Because of the absence of markers that can better discriminate between absolute and functional iron deficiency (e.g., sTfR and/or the sTfR/log ferritin index) or tissue iron staining (e.g., Prussian blue in liver/spleen or bone marrow), our interpretation was rather cautious. This confirmed the need for the inclusion of such markers in future studies to determine whether the iron disturbance during giardiasis is mainly store depletion, inflammatory iron restriction, or a combination of the two (Hanchard and Wonkam, 2021). ConclusionIn this rat model, giardiasis was associated with elevated systemic inflammatory markers and concurrently disrupted iron homeostasis, which included decreased serum iron levels and reduced ferritin levels during acute infection and in the post-clearance phase. These results indicate the possibility of interplay between intestinal infection, inflammation, and iron regulation in weanling rats and warrant further studies of the mechanisms involved using additional biomarkers to differentiate between absolute and functional iron deficiency. This study demonstrates that experimental giardiasis in the rat model was accompanied by inflammation activation, iron regulatory system disruption, and changes in some metabolic indicators. The findings show a triad relationship between intestinal infection, systemic inflammation, and iron metabolism imbalances. The regulation of iron homeostasis and hepcidin synthesis by select metabolites remains speculative and requires further mechanistic studies. AcknowledgmentsThe authors would like to express their sincere appreciation to the College of Veterinary Medicine, University of Al-Qadisiyah, for providing laboratory support and guidance throughout this study. Conflict of interestThe authors have no conflicts of interest to declare. FundingThe authors have self-funded the study. No external funding source is available. Authors’ contributionsZ.F.A.: Investigation, methodology, data curation; writing—original draft. M.A.A.A.: supervision, conceptualization, formal analysis; writing—review & editing; corresponding author. O.H.A.O.: Methodology; bio-plex/assay support; visualization; writing—review & editing. Data availabilityData are available when requested by the corresponding author. ReferencesAbd-Alhassen, J.K., Janabi, A.H.D. and Aboktifa, M.A. 2021. Antioxidant and antimicrobial evaluation of watermelon lycopene. Biochem. Cell. Arch. 21, 2905–2910. Abuga, K.M., Muriuki, J.M., Uyoga, S.M., Mwai, K., Makale, J., Mogire, R.M., Macharia, A.W., Mohammed, S., Muthumbi, E., Mwarumba, S., Mturi, N., Bejon, P., Scott, J.A.G., Nairz, M., Williams, T.N. and Atkinson, S.H. 2022. Hepcidin regulation in Kenyan children with severe malaria and non-typhoidal Salmonella bacteremia. Haematologica 107(7), 1589–1598. Alam, A.M., Ozdemir, C. and Reza, N. 2024. Strongyloides stercoralis in the UK: a systematic review and meta-analysis. Clin. Med. 24(4), 100227. Banerjee, S. and Datta, R. 2023. Localized Leishmania major infection disrupts systemic iron homeostasis that can be controlled by oral iron supplementation. J. Biol. Chem. 299(8), 105064. Barouki, R., Samson, M., Blanc, E.B., Colombo, M., Zucman-Rossi, J., Lazaridis, K.N., Miller, G.W. and Coumoul, X. 2023. The exposome and liver disease—how environmental factors affect liver health. J. Hepatol. 79(2), 492–505; doi: 10.1016/j.jhep.2023.02.034 Bartak, M., Krahel, W.D., Gregorczyk-Zboroch, K., Chodkowski, M., Potârniche, A.V., Długosz, E., Krzyżowska, M. and Cymerys, J. 2025. Cytokine profile analysis during Sialodacryoadenitis virus and mouse hepatitis virus JHM strain infection in primary mixed microglia and astrocyte culture—preliminary research. Cells 14(9), 637; doi:10.3390/cells14090637 Chen, H., Shi, J., Tang, C., Xu, J., Li, B., Wang, J., Zhou, Y., Yang, Y., Yang, H., Huang, Q., Yu, W., Wang, H., Wu, D., Hu, Y., Zhou, H., Sun, Q. and Lu, S. 2024. CHIKV infection drives shifts in the gastrointestinal microbiome and metabolites in rhesus monkeys. Microbiome 12(1), 161; doi: 10.1186/s40168-024-01895-w De Kesel, J., Bonneure, E., Frei, M., De Meyer, T., Mangelinckx, S. and Kyndt, T. 2023. Diproline-induced resistance to parasitic nematodes in rice: roles of iron, nitric oxide and ethylene. Front. Plant Sci. 14, 1112007. Dreischer, P., Duszenko, M., Stein, J. and Wieder, T. 2022. Eryptosis: programmed death of nucleus-free, iron-filled blood cells. Cells 11(3), 503. Frontera, L.S., Moyano, S., Quassollo, G., Lanfredi-Rangel, A., Rópolo, A.S. and Touz, M.C. 2018. Lactoferrin and lactoferricin endocytosis halt Giardia cell growth and prevent infective cyst production. Sci. Rep. 8(1), 18020. Ghazi, A.M., Al-Bayati, M.A. and Janabi, A.H.D. 2024. Metabolomics alterations by phytosomal propolis and lycopene in rats with BPH. Iraqi J. Vet. Sci. 38(Suppl. I–IV), 7–15. Gilabert Carbajo, C., Cornell, L.J., Madbouly, Y., Lai, Z., Yates, P.A., Tinti, M. and Tiengwe, C. 2021. Novel aspects of iron homeostasis in pathogenic bloodstream form Trypanosoma brucei. PLos Pathog. 17(6), e1009696. Gutiérrez, L. and Bartelt, L. 2024. Current understanding of Giardia lamblia and pathogenesis of stunting and cognitive deficits in children from low- and middle-income countries. Curr. Trop. Med. Rep. 11, 28–39; doi:10.1007/s40475-024-00314-2 Hanchard, N.A. and Wonkam, A. 2021. “Iron”ing out hemophagocytosis through PIEZO1. Cell 184(4), 856–858. Kaboré, B., Post, A., Berendsen, M.L.T., Diallo, S., Lompo, P., Derra, K., Rouamba, E., Jacobs, J., Tinto, H., De Mast, Q. and Van Der Ven, A.J. 2020. Red blood cell homeostasis in children and adults with and without asymptomatic malaria infection in Burkina Faso. PLoS One 15(11), 242507. Karim, S.M., Mansour, K.A., Janabi, A.H.D. and Al-Nakeeb, N.K.M. 2018. First phylogenetic characterization of pseudocowpox virus from cattle. Iraqi J. Vet. Sci. 33(1), 123–126. Lozenov, S., Krastev, B., Nikolaev, G., Peshevska-Sekulovska, M., Peruhova, M. and Velikova, T. 2023. Gut microbiome composition regulates malignant transformation, metastasis, and antitumor immunity. Int. J. Mol. Sci. 24(6), 5978. Mach, J. and Sutak, R. 2020. Iron in parasitic protists – from uptake to storage and where we can interfere. Metallomics 12(9), 1335–1347. Moisidis-Tesch, C.M. and Shulman, L.P. 2022. Iron deficiency in women’s health: new insights into diagnosis and treatment. Adv. Ther. 39(6), 2438–2451. Navidifar, T., Meftah, E., Baghsheikhi, H., Kazemzadeh, K., Karimi, H. and Rezaei, N. 2025. Dual role of hepcidin in response to pathogens. Microb. Pathog. 203, 107496. Neves, J.V., Gomes, A.C., Costa, D.M., Barroso, C., Vaulont, S., Cordeiro da Silva, A., Tavares, J. and Rodrigues, P.N.S. 2021. A role for hepcidin in the anemia caused by Trypanosoma brucei infection. Haematologica 106(3), 806–818; doi: 10.3324/haematol.2019.227728 Patel, H., Minkah, N.K., Kumar, S., Zanghi, G., Schepis, A., Goswami, D., Armstrong, J., Abatiyow, B.A., Betz, W., Reynolds, L., Camargo, N., Sheikh, A.A. and Kappe, S.H.I. 2024. Malaria blood stage infection suppresses liver stage infection via host-induced interferons but not hepcidin. Nat. Commun. 15(1), 2104; doi: 10.1038/s41467-024-46270-3 Raj, D., Ghosh, E., Mukherjee, A.K., Nozaki, T. and Ganguly, S. 2014. Differential gene expression in Giardia lamblia under oxidative stress: significance in eukaryotic evolution. Gene 535(2), 131–139. Schytz Andersen-Civil, A.I., Arora, P., Zhu, L., Myhill, L.J., Büdeyri Gökgöz, N., Castro-Mejia, J.L., Leppä, M.M., Hansen, L.H., Lessard-Lord, J., Salminen, J.P., Thamsborg, S.M., Sandris Nielsen, D., Desjardins, Y. and Williams, A.R. 2024. Gut microbiota-mediated polyphenol metabolism is restrained by parasitic whipworm infection and associated with altered immune function in mice. Gut Microbes 16(1), 2370917; doi: 10.1080/19490976.2024.2370917 Sharma, S., Ramachandran, A. and Sharma, A. 2024. Role of riboflavin deficiency in malaria pathophysiology. PLoS Pathog. 20(3), e1011991. Tamarozzi, F., Martello, E., Giorli, G., Fittipaldo, A., Staffolani, S., Montresor, A., Bisoffi, Z. and Buonfrate, D. 2019. Morbidity associated with chronic strongyloides stercoralis infection: a systematic review and meta-analysis. Am. J. Trop. Med. Hyg. 100(6), 1305–1311; doi: 10.4269/ajtmh.18-0895 Woolley, S.D., Grigg, M.J., Marquart, L., Gower, J.S.E., Piera, K., Nair, A.S., Amante, F.M., Rajahram, G.S., William, T., Frazer, D.M., Chalon, S., Mccarthy, J.S., Anstey, N.M. and Barber, B.E. 2024. Longitudinal changes in iron homeostasis in human experimental and clinical malaria. EBioMedicine 105, 105189. Yang, J., Liu, S., Zhao, Q., Li, X. and Jiang, K. 2023. Α-linolenic acid mitigates intestinal inflammation induced by Toxoplasma gondii. Microbiome 11(1), 273. Yaseen, M.M., Karawan, A.C., Alfatlawi, M.A.A. and Janabi, A.H.D. 2020. Gut bacterial cytochrome-P450 in mosquito larvae degrades temephos insecticide. Ann. Trop. Med. Public Health 23(1), S412. Zakiyah, Z., Jayanti, Y.D., Nurdiana, N. and Dwijayasa, P.M. 2017. Effects of time course ferrous sulphate supplementation on iron regulation in pregnant rats. J. Taibah Univ. Med. Sci. 12(2), 146–149; doi:10.1016/j.jtumed.2016.12.007 | ||
| How to Cite this Article |
| Pubmed Style Abbas ZF, Al-obaidy OH, Alfatlawi MAA. Effects of giardiasis on iron, hepcidin, and gut microbiota metabolites in young rats: Evidence for systemic inflammation and malabsorptive metabolic reprogramming. Open Vet. J.. 2026; 16(5): 3104-3120. doi:10.5455/OVJ.2026.v16.i5.52 Web Style Abbas ZF, Al-obaidy OH, Alfatlawi MAA. Effects of giardiasis on iron, hepcidin, and gut microbiota metabolites in young rats: Evidence for systemic inflammation and malabsorptive metabolic reprogramming. https://www.openveterinaryjournal.com/?mno=298577 [Access: June 26, 2026]. doi:10.5455/OVJ.2026.v16.i5.52 AMA (American Medical Association) Style Abbas ZF, Al-obaidy OH, Alfatlawi MAA. Effects of giardiasis on iron, hepcidin, and gut microbiota metabolites in young rats: Evidence for systemic inflammation and malabsorptive metabolic reprogramming. Open Vet. J.. 2026; 16(5): 3104-3120. doi:10.5455/OVJ.2026.v16.i5.52 Vancouver/ICMJE Style Abbas ZF, Al-obaidy OH, Alfatlawi MAA. Effects of giardiasis on iron, hepcidin, and gut microbiota metabolites in young rats: Evidence for systemic inflammation and malabsorptive metabolic reprogramming. Open Vet. J.. (2026), [cited June 26, 2026]; 16(5): 3104-3120. doi:10.5455/OVJ.2026.v16.i5.52 Harvard Style Abbas, Z. F., Al-obaidy, . O. H. & Alfatlawi, . M. A. A. (2026) Effects of giardiasis on iron, hepcidin, and gut microbiota metabolites in young rats: Evidence for systemic inflammation and malabsorptive metabolic reprogramming. Open Vet. J., 16 (5), 3104-3120. doi:10.5455/OVJ.2026.v16.i5.52 Turabian Style Abbas, Zahraa Fadhil, Osama Hadi Al-obaidy, and Monyer Abdulameir Abd Alfatlawi. 2026. Effects of giardiasis on iron, hepcidin, and gut microbiota metabolites in young rats: Evidence for systemic inflammation and malabsorptive metabolic reprogramming. Open Veterinary Journal, 16 (5), 3104-3120. doi:10.5455/OVJ.2026.v16.i5.52 Chicago Style Abbas, Zahraa Fadhil, Osama Hadi Al-obaidy, and Monyer Abdulameir Abd Alfatlawi. "Effects of giardiasis on iron, hepcidin, and gut microbiota metabolites in young rats: Evidence for systemic inflammation and malabsorptive metabolic reprogramming." Open Veterinary Journal 16 (2026), 3104-3120. doi:10.5455/OVJ.2026.v16.i5.52 MLA (The Modern Language Association) Style Abbas, Zahraa Fadhil, Osama Hadi Al-obaidy, and Monyer Abdulameir Abd Alfatlawi. "Effects of giardiasis on iron, hepcidin, and gut microbiota metabolites in young rats: Evidence for systemic inflammation and malabsorptive metabolic reprogramming." Open Veterinary Journal 16.5 (2026), 3104-3120. Print. doi:10.5455/OVJ.2026.v16.i5.52 APA (American Psychological Association) Style Abbas, Z. F., Al-obaidy, . O. H. & Alfatlawi, . M. A. A. (2026) Effects of giardiasis on iron, hepcidin, and gut microbiota metabolites in young rats: Evidence for systemic inflammation and malabsorptive metabolic reprogramming. Open Veterinary Journal, 16 (5), 3104-3120. doi:10.5455/OVJ.2026.v16.i5.52 |

