Open Veterinary Journal, (2026), Vol. 16(5): 3080-3093
Research Article
10.5455/OVJ.2026.v16.i5.50
Anti-Cryptosporidium efficacy and metabolite profiling of Schefflera arboricola L. leaves extract
Amal M. El-Feky1, Amany M. Mohamed2, Magdy A. Ayad3*, Nehal M. Khairy4 and
Raafat Mohamed Shaapan5
1Pharmacognosy Department, Pharmaceutical and Drug Industries Research Institute, National Research Centre, Giza, Egypt
2Parasitology and Animal Diseases Department, Veterinary Research Institute, National Research Centre, Giza, Egypt
3Department of Microbiology and Parasitology, Faculty of Veterinary Medicine, University of Tripoli, Tripoli, Libya
4Department of Microbiology and Immunology, Egypt Drug Authority (EDA), (Formerly NODCAR), Giza, Egypt
5Zoonotic Diseases Department, Veterinary Research Institute, National Research Centre, Giza, Egypt
*Corresponding Author: Magdy A. Ayad. Department of Microbiology and Parasitology, Faculty of Veterinary Medicine, University of Tripoli, Tripoli, Libya. Email: magdyaa73 [at] yahoo.com
Submitted: 19/11/2025 Revised: 14/03/2026 Accepted: 24/03/2026 Published: 31/05/2026
© 2025 Open Veterinary Journal
This is an Open Access article distributed under the terms of the Creative Commons Attribution-Non Commercial-No Derivatives License (http://creativecommons.org/licenses/by-nc-nd/4.0/), which permits non-commercial re-use, distribution, and reproduction in any medium, provided the original work is properly cited, and is not altered, transformed, or built upon in any way.
Abstract
Background: Schefflera arboricola L., (S. arboricola L) is a member of the Araliaceae family and exhibits various biological activities. Cryptosporidium parvum, a zoonotic protozoan infection, can infect a variety of vertebrate and invertebrate hosts, including humans.
Aim: This study aimed to explore the phytochemical constituents and efficacy of S. arboricola leaf extracts against C. parvum infection.
Methods: The assessment of chlorophyll, carotenoid, carbohydrate, total phenolic, and flavonoid contents in the leaves of S. arboricola was quantified using high-performance liquid chromatography and UPLC/ESI–MS. Parasitological, histological, deoxyribonucleic acid (DNA) fragmentation, liver functions, lipid profiles, and antioxidant parameter analysis will be used to assess the efficacy of the treatments after oral infection with C. parvum oocytes (2 × 103/mouse). Fifty adult male mice were divided into five groups (10 each): non-treated Cryptosporidium-infected group (G1), infected and treated with low-dose S. arboricola extract (100 mg/kg) (G2), infected and treated with high-dose S. arboricola extract (200 mg/kg) (G3), Nitazoxanide treated (150 mg/kg) (G4), and non-infected group (G5).
Results: This study quantified the total phenolic and flavonoid contents 267.54 ± 0.61 mg GAE/g and 514.89 ± 0.20 mg rutin/g, respectively, and identified 51 compounds, including 14 phenolic acids and their derivatives, 30 flavonoids, 6 coumarins, and 1 anthocyanin. Both infected-treated mice groups (G2 & G3) showed a considerable improvement in Cryptosporidium oocyte shedding, with significant effectiveness (p < 0.05). In addition, the infected-treated groups showed less severe changes in the intestinal epithelium structures and a significant decrease in DNA fragmentation, hepatic biomarkers, and lipid indices compared with the infected non-treated control group.
Conclusion: Schefflera arboricola leaves contain a wide variety of pigments, carotenoid, carbohydrate, phenolic, and flavonoid compounds. Extracts of S. arboricola leaves are a new interesting prospective natural product that may be used in the treatment of Cryptosporidium infection.
Keywords: Schefflera arboricola L, Phytochemical, Natural product, Anti-Cryptosporidium.
Introduction
Schefflera arboricola L. (S. arboricola L) represents the most extensive genus within the Araliaceae family, encompassing more than 650 species found in tropical and subtropical regions. Various species have a documented history of traditional medicinal applications in China, India, and Indonesia (Wang et al., 2020). Schefflera arboricola, which belongs to the Araliaceae family, is indigenous to Taiwan. Commonly referred to as the "Dwarf umbrella tree" (Plunkett et al., 2005), this evergreen shrub typically reaches heights of 3–4 m. Its leaves are palmately compound, featuring 7–9 leaflets that measure 10–20 cm in length and 4–6 cm in width (Zhao et al., 2010). This ornamental plant is widely regarded as a favored choice for indoor gardening due to its adaptability to various unfavorable growing conditions (Wang et al., 2021).
The S. arboricola plant and its extracts have a long-standing history of possessing strong anti-inflammatory and analgesic effects, so they are utilized for alleviating stomach pain, headaches, trigeminal neuralgia, rheumatic arthritis, fractures, and sprains (Chang and Butt, 1986; Zhao et al., 2010; Liu et al., 2011). The leaves and bark of Schefflera are recognized for their antioxidant properties and are used as a treatment for coughs and as a diuretic (Deepa and Nalini, 2013). The phytochemical composition of S. arboricola leaves and stems has resulted in the isolation of two oleanolic acid glucuronides, specifically cynarasaponin H, olaxoside, lupine, triterpene, and saponins (Melek et al., 2003; Guo et al., 2006). Furthermore, S. arboricola L. leaves are thought to be a significant source of bioactive phenolic substances; Gas Chromatography-Mass Spectrometry and Liquid Chromatography–Electrospray Ionization–Mass Spectrometry (LC-ESI-MS) analysis revealed the presence of 29 phenolic compounds (El-Hagrassi et al., 2022).
The apicomplexan parasite Cryptosporidium spp. mostly affects the gastrointestinal tracts of humans and animals. The infection is common in calves and children and can cause severe diarrhea, anorexia, and dehydration (Ghazy et al., 2015a; Lombardelli et al., 2019). Cryptosporidiosis is a zoonotic parasite that can spread from infected animals to humans. Transmission occurs through the ingestion of oocytes, through either direct contact or consumption of contaminated water or food (Toaleb and Shaapan, 2024). There is growing evidence that a variety of wildlife species, including fish, amphibians, reptiles, birds, and mammals, are infected with cryptosporidiosis (Saad-Alla et al., 2022; Shaapan et al., 2022). A panel of experts from the Food and Agriculture Organization and the World Health Organization have determined that Cryptosporidium is the fifth most important food-borne parasite in the world (Javed and Alkheraije, 2023).
The only treatment currently approved by the Food and Drug Administration (FDA) to treat Cryptosporidium infections in humans, particularly immunocompetent patients, is nitazoxanide (NTZ), which decreases oocyte shedding and shortens the severity of diarrhea (Shoukry et al., 2025). In addition to NTZ, a few other commercially available medications, such as paromomycin and azithromycin, have some anti-Cryptosporidium efficacy, but the FDA has not licensed them to treat cryptosporidiosis (Caravedo and White, 2023). Consequently, the development of alternative, plant-based therapeutics with improved efficacy and safety profiles has become a research priority. Traditional plant extracts, such as onion (Allium cepa) and cinnamon (Cinnamomum zeylanicum) oils (Abu El Ezz et al., 2011), ginger (Zingiber officinale), ginseng (Panax ginseng), and sage (Salvia officinalis) (Abouelsoued et al., 2020), and Citrus sinensis peel extract (Abd El Wahab et al., 2022).
Despite the extensive traditional use and recognized pharmacological potential of S. arboricola, comprehensive studies on its phytochemical composition, particularly detailed metabolite profiling of phenolic and flavonoid constituents, remain scarce. Moreover, no previous research has evaluated its efficacy against Cryptosporidium parvum infection. Therefore, the primary aim of the present study was to perform an in-depth phytochemical analysis of S. arboricola leaves and evaluate the anti-cryptosporidial efficacy of its extract in experimentally infected mice.
Materials and Methods
Chemicals
The chemicals and solvents used in this investigation were of analytical grade and were supplied by Merck and Co., Inc. (USA) and Sigma-Aldrich (USA). The high-performance liquid chromatography (HPLC) specifications and all tested compounds met the highest analytical purity criteria. Phenolic compounds and flavonoids analyzed in this study were acquired from Sigma-Aldrich Co. (USA).
Plant collection and extraction
Uniform young leaves of S. arboricola L. were collected from Orman Garden, Giza, Egypt in October 2024. Mrs. Trease Labib, a Plant Taxonomy Consultant at the Ministry of Agriculture, identified and authenticated the plant. A voucher specimen (Voucher No. M242) was deposited in the herbarium of the National Research Center, Cairo, Egypt, for future reference. The collected leaves were air-dried in the shade at room temperature to prevent the degradation of thermolabile constituents and then ground into a fine powder. A total of 400 g of the powdered leaves was subjected to cold maceration with absolute methanol (1.5 L × 6) to ensure exhaustive extraction of both polar and semipolar metabolites were extracted.
Detection of pigments and carbohydrates
The pigment concentration was quantified using spectrophotometry. Chlorophyll a and chlorophyll b concentrations were measured at wavelengths of 651 and 664 nm, respectively, while total carotenoids were evaluated at a wavelength of 470 nm. The calculations for total chlorophylls, total carotenoids, chlorophyll ratio (a/b), and pigment index (the ratio of total carotenoids to chlorophyll a) were based on the formulas provided by Bodnar et al. (2016). Furthermore, the total carbohydrate content was quantitatively determined as glucose using the phenol sulfuric acid method in accordance with Dubois et al. (1956).
Total phenolic and flavonoid detection
The Folin–Ciocalteu method was used to determine the total phenolic content in the methanol extract of S. arboricola L. leaves, as reported by El-Feky et al. (2017). The results were expressed in terms of the gallic acid equivalent. The total flavonoid content was evaluated using the aluminum chloride method, as described by Baba and Malik (2015), with the flavonoid concentration represented as rutin equivalent. All results were presented as mean ± S.D.
Animals
Fifty adult male mice with an average body weight of 25–30 g were housed in the Laboratory Animal House at the National Research Center in Giza, Egypt. Mice are given a regular laboratory meal and unlimited access to water while being kept as 10 mice per cage with 12:12 hour day/night cycles at a temperature of (20ᵒC–25ᵒC) (Shaapan et al., 2023).
Design of the experiments and infection
Mice were divided into five groups (10 each): Cryptosporidium-infected non-treated group (G1), infected and treated with a low dose of S. arboricola extract (100 mg/kg) (G2), infected and treated with a high dose of S. arboricola extract (200 mg/kg) (G3), infected and treated with NTZ (150 mg/kg) (G4), and non-infected non-treated group (G5). Oocytes from a stock of Cryptosporidium spp. that was kept in the Zoonotic Disease Department, National Research Centre, were genetically identified as C. parvum (Hassanain et al., 2011). Mice infected by a single oral dosage of 2 × 103 C. parvum oocytes for each mouse using oral-gastric gavage treatment with S. arboricola extract and NTZ medications will start on the third day post-infection, and all tested mice will be sacrificed 3 weeks post-infection (Abouelsoued et al., 2023).
Parasitological examination
Throughout the experiment, fecal samples from each group were separately collected every day, and the amount of Cryptosporidium oocytes was counted using MZN stain (Ghazy et al., 2016). Centrifugation of the fecal suspension for 10 minutes at 3,000 rpm. 100 μl of fecal sediment was used to measure the number of oocytes in 1 ml of fecal sample, which was then observed via an oil immersion lens (×100). The average of three computed counts, multiplied by 10, and the number of oocytes per gram of feces were stated (Elfadaly et al., 2018).
Histopathological examination
The intestinal portion (ileum) of each mouse group was preserved in a 10% buffered formalin solution before being embedded in blocks of paraffin wax. Hematoxylin and eosin staining, sectioning, and light microscopy examination (Barakat et al., 2024).
Quantification of deoxyribonucleic acid (DNA) fragmentation
The DNA fragmentation of the tested mice was detected in the intestinal epithelium using the PEG/Hoechst fragmentation assay. This technique is based on the idea that centrifugal sedimentation can be used to separate broken-up double-stranded DNA from chromosomic DNA. High-molecular-weight DNA was preferentially precipitated using polyethylene glycol after cells were broken down with hypotonic acid. The fluorescent dye Hoechst 33258 was used in a fluorometer to measure the amount of fragmented DNA remaining in the supernatant (Loannou and Chen, 1996; Shaapan et al., 2011).
Biochemical marker estimation
The sera collected from all examined groups of mice were analyzed for lipid profile: total cholesterol (TC), triglycerides (TG), high-density lipoprotein (HDL), and low-density lipoprotein (LDL). Sera liver function enzymes: aspartate aminotransferase (AST) and alanine aminotransferase (ALT). In addition, the oxidative stress marker malondialdehyde (MDA) was also estimated in the sera. The biochemical analysis was carried out in accordance with Lütjohann et al. (2023), and the test kits were supplied by Bio-diagnostic, Egypt, and BioMérieux-France.
Statistical analysis
A one-way analysis of variance (one-way ANOVA) was performed using the Statistical Package for Social Sciences (SPSS for Windows, version 11.0) to evaluate the positive and negative statistical relationships among different biological findings.
Ethical approval
The study was approved by the ethical review board of the Medical Research Ethics Committee at the National Research Centre in Egypt. The ethical approval certificate was documented under the registration number: (0441 in 29/04/2025).
Results
Content of pigments and carbohydrates
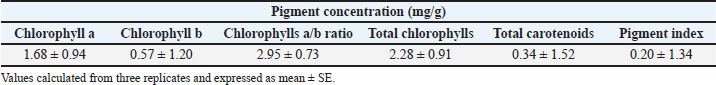
The chlorophyll and carotenoid levels in the leaves of S. arboricola L. were evaluated, and the results demonstrated that the chlorophyll a and b concentrations were 1.68 and 0.57 mg/g, respectively. The chlorophyll a/b ratio was calculated as 2.95 mg/g. The total chlorophyll content was 2.28 mg/g, while the total carotenoid content was 0.34 mg/g. The pigment index of the leaves was recorded at 0.20 mg/g. The total carbohydrate content in the leaves of S. arboricola L. was 23.5% (wt/wt), represented as glucose (Table 1).
Table 1. Pigment contents in S. arboricola L. leaves.
Phenolic and flavonoid contents
The total phenolics and flavonoids were quantified in the methanol extract from the leaves of S. arboricola L., resulting in measurements of 267.54 ± 0.61 mg GAE/g and 514.89 ± 0.20 mg rutin/g, respectively. These findings affirm the considerable presence of total phenolics and flavonoids in the leaves of S. arboricola L., which serves as a basis for a diverse array of biological functions.
The methanol extract from the leaves of S. arboricola L. was subjected to HPLC to ascertain the presence of various phenolic acids and flavonoids, as shown in Table 2 and Figure 1. The results of the analysis identified 19 distinct compounds, with rosmarinic acid being the predominant one, recorded at a retention time of 11.830 and a concentration of 17,926.41 µg/g. Other notable compounds included quercetin, caffeic acid, naringenin, gallic acid, kaempferol, and hesperetin, with respective concentrations of 2,657.63, 2,071.95, 1,946.32, 1,531.69, 1,061.24, and 1,052.41 µg/g.
Table 2. HPLC identification of polyphenolic compounds in the methanol extract from S. arboricola L.
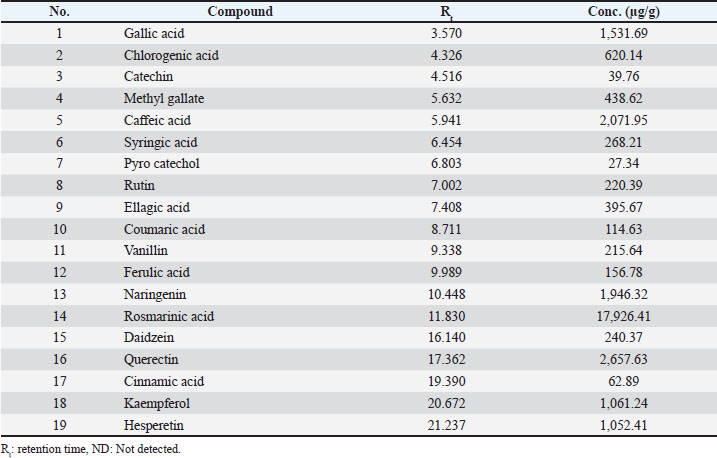
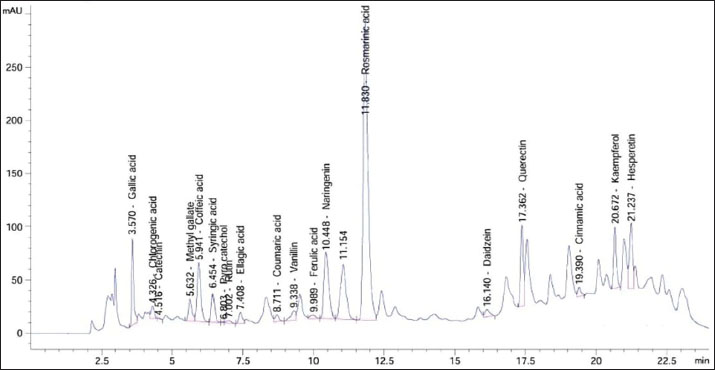
Fig. 1. High-performance liquid chromatography (HPLC) chromatogram for polyphenolic compounds in the methanol extract from S. arboricola L.
Furthermore, the Ultra-Performance Liquid Chromatography–Electrospray Ionization–Mass Spectrometry (UPLC/ESI–MS) analysis performed in both negative and positive ionization modes to thoroughly characterize the diverse phytochemical classes found in the leaves of S. arboricola L. This investigation revealed the existence of 51 compounds, including 14 phenolic acids and their derivatives, 30 flavonoids, 6 coumarins, and 1 anthocyanin (Fig. 2).
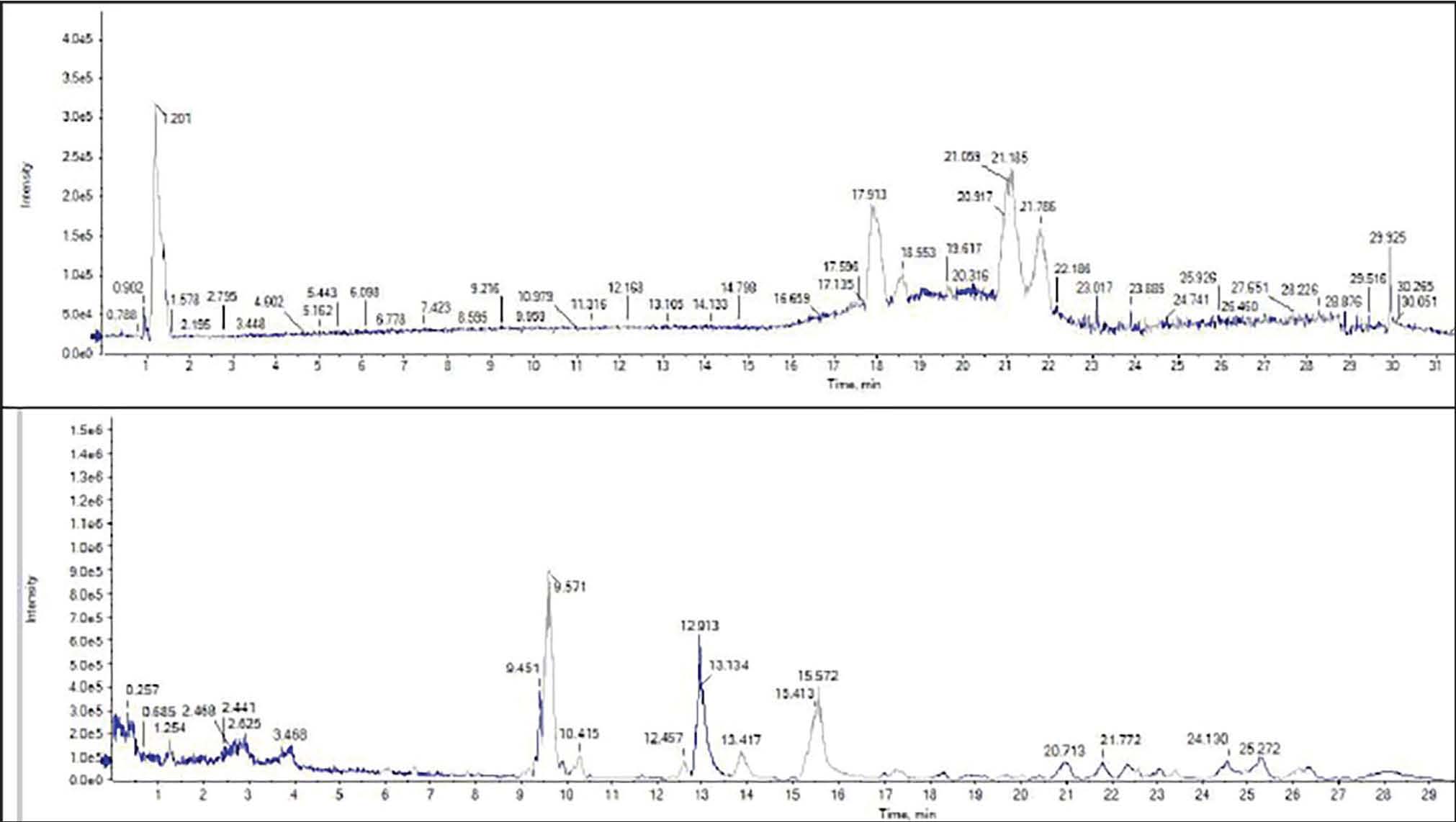
Fig. 2. UPLC/ESI–MS chromatogram for the polyphenolic profile in the methanol extract from S. arboricola L.
Morphological identification of Cryptosporidium oocytes
The obtained mZN-stained C. parvum oocytes shed in the feces of experimentally infected mice showed a smooth wall, ovoid to spherical-shaped oocytes, and occasionally showed an incomplete suture line at the oocytes wall and presented as red-pink (acid fast) with green-back (Fig. 3).
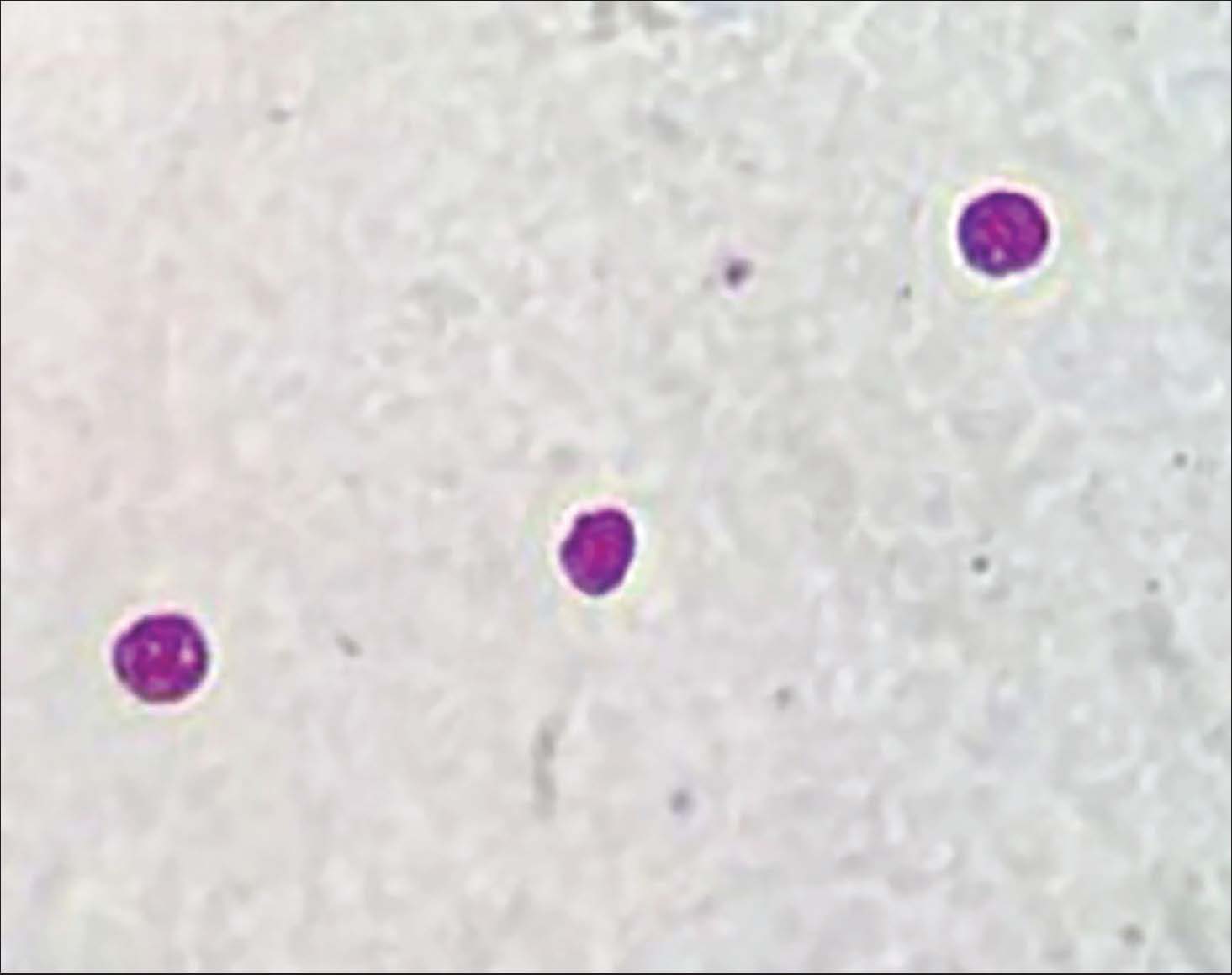
Fig. 3. Cryptosporidium oocytes in mice fecal smears stained with Modified Ziehl-Neelsen (MZN) (x400).
Cryptosporidium oocyte shedding
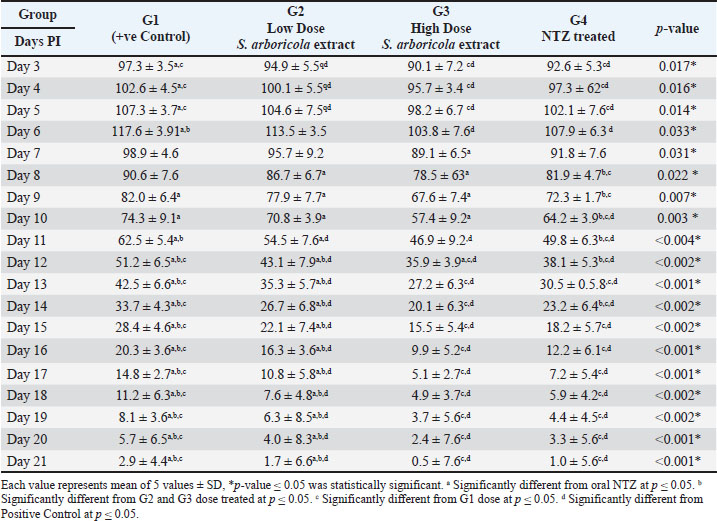
The count of Cryptosporidium oocytes was significantly higher in the fecal samples of the infected non-treated (positive control) group (G1) than in the other treated infected groups during all experimental periods. Oocyte shedding gradually decreased until few oocytes were detected on day 7 PI. In the groups treated with S. arboricola L. extract (G2 & G3) and NTZ (G4), there was a statistically significant decrease in oocyte shedding. Additionally, the higher-dose S. arboricola L (200 mg/kg)-treated group (G3) revealed decreased oocyte shedding throughout time, followed by the NTZ-treated group (G4), whereas the low-dose S. arboricola L (100 mg/kg)-treated group (G2) showed the highest oocyte shedding than the other two treatment groups (Table 3).
Table 3. Cryptosporidium oocytes in mice fecal smears stained with Modified Ziehl-Neelsen (MZN) (x400).
Histopathological investigation
The intestinal epithelial mucosa of each mouse group had a markedly different look at the end of the experiment (PI, 3 weeks). The infected non-treated mice (positive control) (Fig. 4A) exhibited prominent histopathological changes in the intestinal mucosa, including enlarged, shortened, and desquamated intestinal villi, indicating a significant degree of villous atrophy with chronic inflammatory cell infiltration in the lamina propria. Conversely, the histological alterations were more pronounced in the other treatment groups (Fig. 4 B–D), and there was evidence of a higher number of goblet cells and a modest degree of villous atrophy with little inflammatory cell and modest edema in the lamina propria. Mice that were neither infected nor receiving treatment (negative control) (Fig. 4E) had normal intestinal mucosa morphology, with several finger-like villi and without any pathological changes in the mucosa or lamina propria. Additionally, the histopathological changes improved more in high-dose (200 mg/kg) (Fig. 4C) than low-dose (100 mg/kg) S. arboricola L-treated mice (Fig 4B).
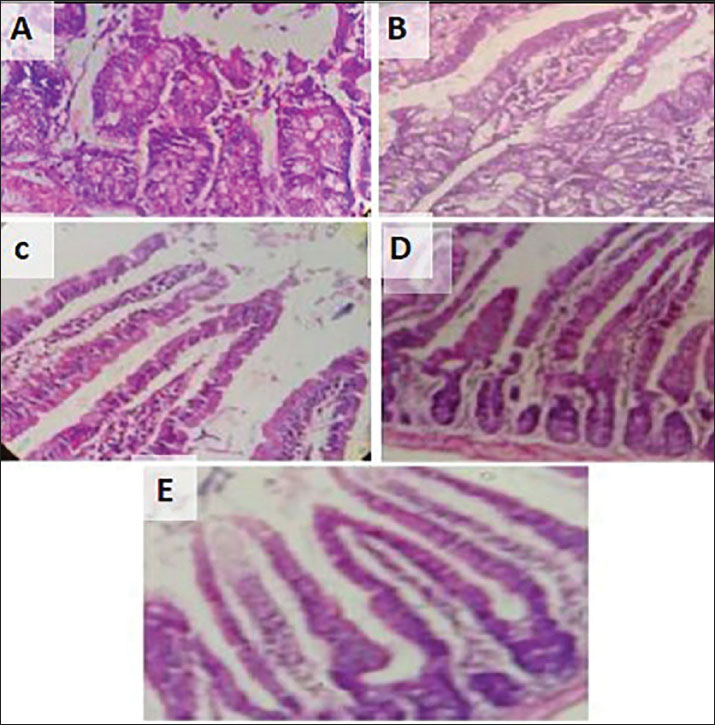
Fig. 4. Mice intestine; C. parvum infected nonrated (A), infected low dose treated with plant extract (B), infected treated with high dose of plant extract (C), infected treated with NTZ (D), and non-infected (E).
DNA fragmentation
The high-dose S. arboricola L-treated mice showed a decline in the DNA fragmentation quantitative results (54.47), followed by NTZ-treated (74.92) and low-dose S. arboricola L-treated (103.95) mice compared with the infected non-treated mice (209.03) (Fig. 5).
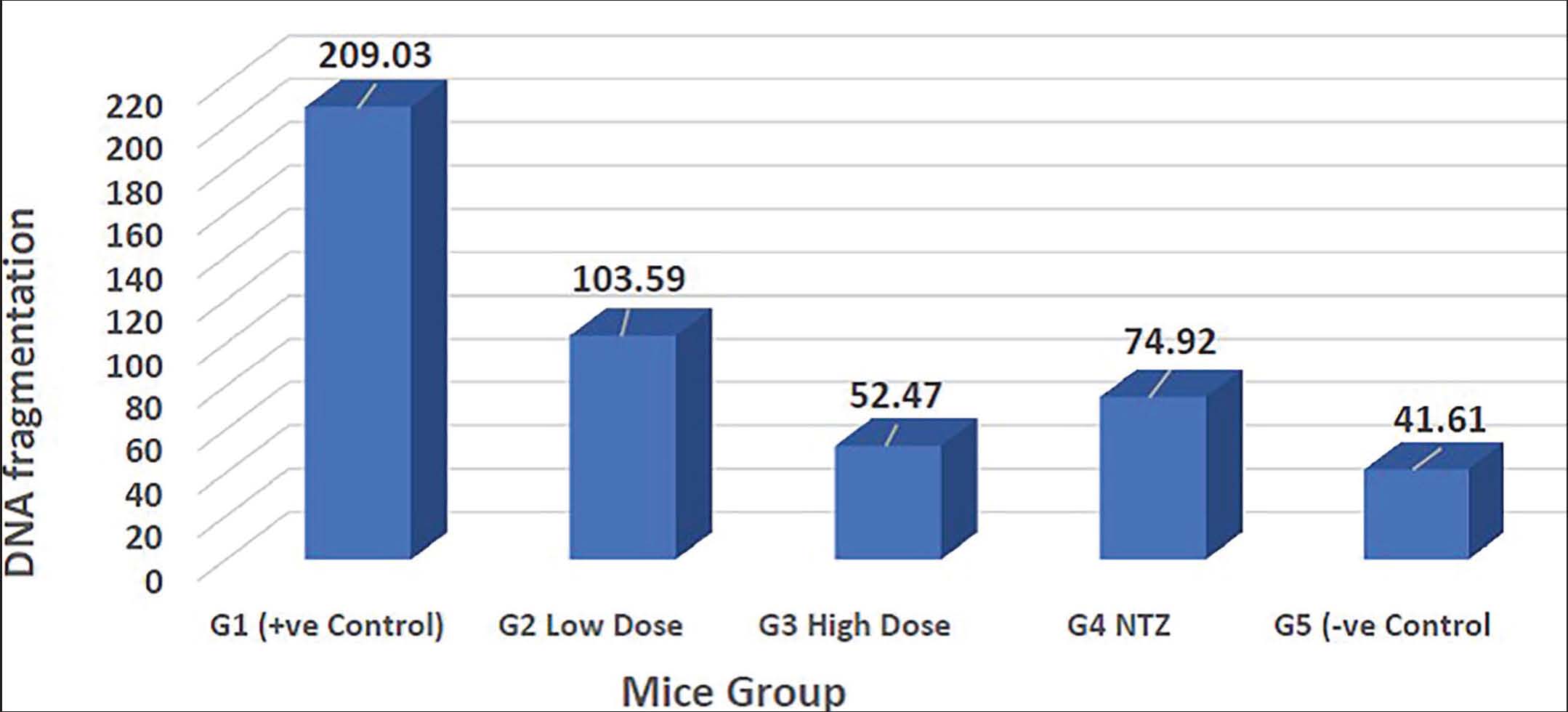
Fig. 5. DNA fragmentation in Cryptosporidium experimentally infected non-treated (G1), infected treated with low-dose S. arboricola (G2), infected treated with high-dose S. arboricola (G3), infected treated with NTZ (G4), and non-infected non-treated (G5).
Biochemical parameters
The results showed that C. parvum-infected non-treated mice (G1) had a higher lipid profile (TC, TG, HDL, and LDL), liver enzymes (ALT and AST), and oxidative stress marker MDA compared to non-infected mice (G5). Meanwhile, the groups infected with S. arboricola and NTZ (G2, G3, and G4) demonstrated a significant decline in all biochemical parameters (Fig. 6).
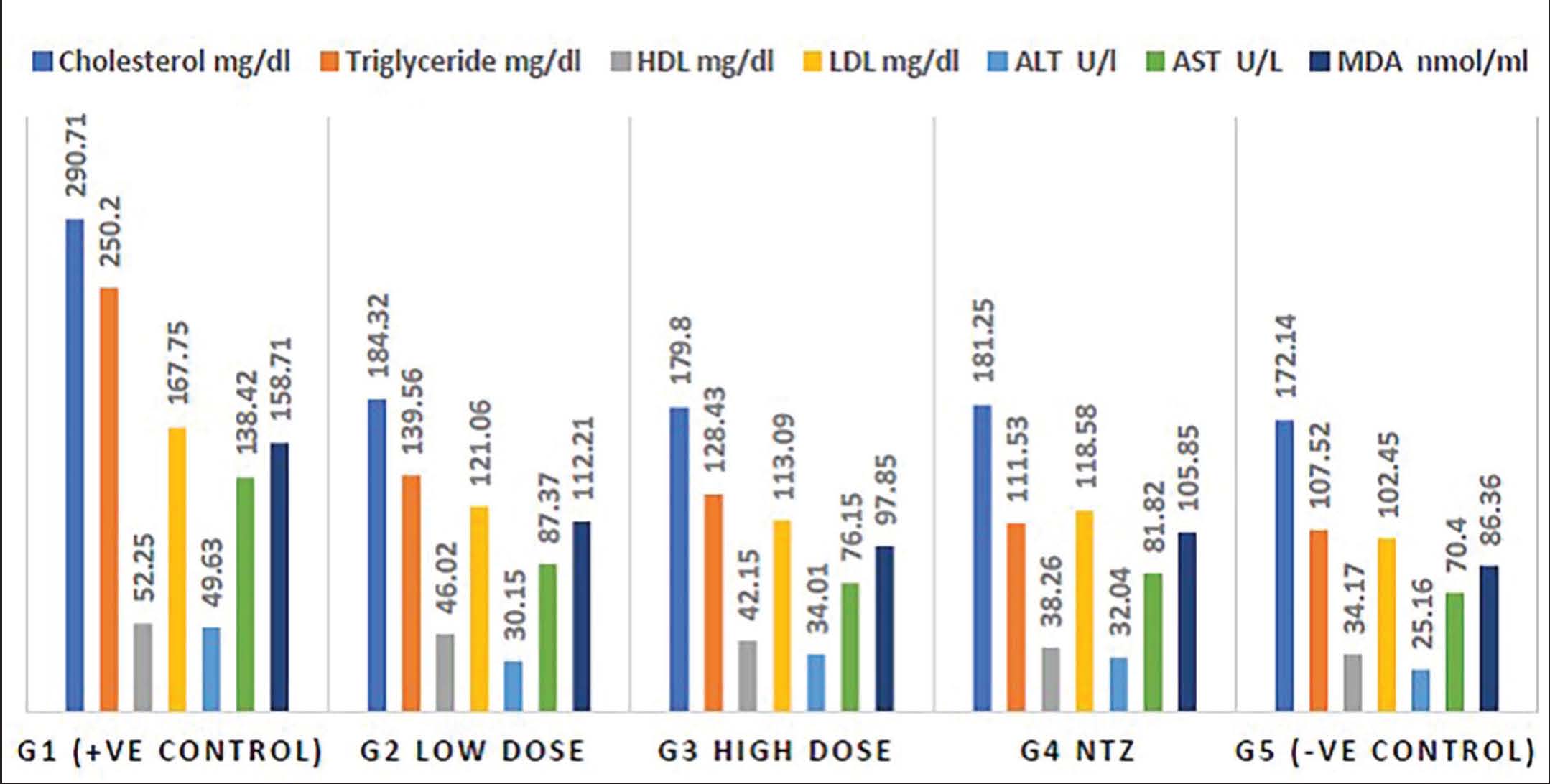
Fig. 6. Biochemical parameters in mice Cryptosporidium experimentally infected non-treated (G1), infected treated with low-dose S. arboricola (G2), infected treated with high-dose S. arboricola (G3), infected treated with NTZ (G4), and non-infected non-treated (G5).
Discussion
In the current study, LC-ESI-MS analysis of S. arboricola leaf extract revealed a complex profile of phenolic acids and their derivatives. These compounds are characterized by high polarity, which accounts for their early elution during CS. Most phenolic acids were identified in the negative ionization mode, with limited detection in the positive mode, corresponding to compounds 1–6, 8–14, and 37, respectively. According to Ristivojević et al. (2015), the fragmentation behavior of phenolic acids typically involves the release of CO₂ and H₂O groups, a pattern also observed in this study. For instance, ferulic acid (compound 1, Rt 1.05) displayed a molecular ion [M–H]⁻ at m/z 193.989 and a fragment ion at m/z 178, resulting from the loss of a methyl group (Gardana et al., 2007). Similarly, compound 9 exhibited precursor ions at m/z 353.007 (negative mode) and 355.103 (positive mode), consistent with that of caffeoylquinic acid (a chlorogenic acid derivative). Its MS/MS fragmentation pattern was characterized by ester bond cleavage between caffeoyl and quinic acid moieties, dehydration of quinic acid to m/z 173 [quinic acid–H–H₂O], and decarboxylation of caffeoyl yielding m/z 135 [caffeoyl–H–CO₂] (Ruan et al., 2019). Compound 12, identified as dicaffeoylquinic acid, appeared in the negative mode at Rt 4.84 with a molecular ion [M–H]⁻ at m/z 515.176, producing fragment ions at m/z 353, 191, and 179, reflecting successive caffeoyl losses and quinic dehydration (Ruan et al., 2019).
In addition to phenolic acids, the LC-ESI-MS profiling of S. arboricola leaves revealed a rich diversity of flavonoid subclasses in both negative and positive ionization modes. The identified compounds included six flavonoid aglycones, 14 flavonoid monoglycosides, 5 flavonoid diglycosides, 4 methoxy flavonoids, and a single bioflavonoid, diosmin (compound 43). These findings are consistent with those of previous reports (El-Hagrassi et al., 2022), confirming the wide spectrum of phenolic and flavonoid constituents in this species. Methoxy flavones exhibit characteristic methyl loss during fragmentation, with m/z values of 298, 330, 358, and 328, respectively (Pellati et al., 2011). A detailed examination of the negative ionization mode spectra revealed that quercetin (compound 42), kaempferol (compound 46), and isorhamnetin (compound 47) followed the classical Recommended Dietary Allowance fragmentation pathway, generating dominant [1,3A]- and [1,2A]- fragments at m/z 179 and 151, respectively (Fabre et al., 2001). Flavonoid glycosides identified through the cleavage of glycosidic O-linkages in both ionization modes, leading to the loss of monosaccharide residues and H-rearrangement. In compounds 21, 29, 34, 39, 43, 48, and 50, the removal of rutinoside (m/z 308 amu) yielded aglycone-specific fragments (Kečkeš et al., 2013). Similarly, compounds 16, 17, 18, 20, 22, 28, and 41 underwent fragmentation through the loss of hexosides (glucose or galactose, m/z 162 amu), whereas compounds 35 and 36 eliminated rhamnose (m/z 146 amu). Compounds 38 and 45 (arabinose or xylose, m/z 132 amu) were lost (Hossain et al., 2010). Characteristic fragmentation of C-glycosides involves dehydration and (0.2)/(0.3)-ring cleavages with elimination of ions at m/z 120 and 150, a hallmark of C-linkages (Farag et al., 2016). In addition to flavonoids, six coumarins—esculetin, 6,7-dimethoxycoumarin, scopoletin, and esculin—were identified in both ionization modes. For example, scopoletin (compound 23, Rt 11.95) displayed a molecular weight of 193.158 in positive mode and produced fragment ions at m/z 178 (loss of CH3), 149 (loss of CO), and 133 (loss of CH3COOH) (Zeng et al., 2015).
Using a modified Ziehl-Neelsen stain, we discovered that Cryptosporidium oocytes in the mucosa of intestinally scraped mice had a smooth, circular, bright red wall. Identification based on standards, such as oocyte size and morphology (Xiao et al., 2004). Cryptosporidium parvum mostly infects the gastrointestinal epithelium of humans and animals. It lives at the apical portion of the epithelial cells and does not cause systemic infection or deep tissue penetration (Ghazy et al., 2015b). The ileum was chosen for this investigation because it is the region most affected by cryptosporidiosis in mice. Cryptosporidiosis severity depends on the level of immunosuppression (Oliveira et al., 2019). According to the parasitological results of the investigation, the fecal samples of the infected non-treated group (positive control) had significantly higher levels of Cryptosporidium oocytes shedding than those of the other Moringa oleifera-treated infected groups across all experimental periods (Ghazy et al., 2016). This finding, as well as the fact that the infected non-treated mice exhibited fewer oocytes shedding and pathological lesions compared to the infected treated groups, was noted in oral NTZ-treated groups on days 6–15 following infection (Abouelsoued et al., 2020). These modifications to the histopathological appearance of Cryptosporidium-infected mice are in line with previous research. The intestinal mucosa of the untreated infected mice showed obvious histopathological changes, such as shortened and widened intestinal villi. This indicated a significant level of villous atrophy and desquamation of the epithelial lining layer in the majority of the villi (Abu El Ezz et al., 2011; Abd El Wahab et al., 2022). Cryptosporidiosis management is difficult due to the low infectious dosage, the enormous number of sporulated oocytes that shed from different hosts, and the stability of the oocytes in the environment. Certain drugs, such as lasalocid, nitazoxanide, aminoglycoside, and dinitroanilineoryzalin, have been shown to reduce the number of Cryptosporidium oocytes in vitro (Innes et al., 2020). There are numerous explanations for why the intestinal disorders and total quantity of Cryptosporidium oocytes in mice given S. arboricola may have strong potential antioxidant, immunomodulatory, and anti-inflammatory effects, which could account for the study’s notably beneficial and corrective outcomes (Abouelsoued et al., 2023). This could account for S. arboricola’s apparent anti-Cryptosporidium action, particularly at higher doses (200 mg/kg).
The observed anti-Cryptosporidium activity of S. arboricola leaf extract is most likely due to its rich phenolic and flavonoid composition rather than a single bioactive constituent. Phenolic acids such as rosmarinic acid, one of the predominant compounds identified in the extract, have demonstrated antiparasitic and antimicrobial properties and represent promising scaffolds for development against protozoan targets, including Giardia species, through enzyme inhibition and oxidative stress mechanisms. (Kyriakopoulou et al., 2025). Flavonoids such as quercetin and kaempferol have been reported to exert antiparasitic effects against a range of protozoa. In vitro studies have shown that certain flavonoids—including quercetin and naringenin—display activity against C. parvum, supporting their relevance to the anti-Cryptosporidium activity observed in this study (Mead and McNair, 2006). In addition, quercetin and kaempferol are known for their broad antiprotozoal activity against other gastrointestinal parasites, such as Giardia lamblia, Leishmania spp., and entameba spp., frequently acting through mechanisms involving disruption of parasite metabolism, oxidative damage, or interference with enzyme systems essential for survival. Caffeic acid and chlorogenic acid derivatives—also identified in the extract—have shown antiparasitic effects in vitro, including growth inhibition of Giardia trophozoites, likely via reactive oxygen species induction and parasite energy metabolism disruption. Although flavanones such as naringenin and hesperetin often exhibit weaker direct anti-parasitic activity, they may contribute through antioxidant and host immunomodulatory effects that enhance defense mechanisms against infection (Alday-Provencio et al., 2015). Collectively, these findings suggest that the anti-Cryptosporidium activity of S. arboricola leaf extract arises from the synergistic action of multiple phenolic acids and flavonoids, consistent with previous reports on plant-derived antiparasitic extracts.
The biochemical findings in the present study demonstrated that C. parvum infection induced significant alterations in hepatic function markers, lipid profile, and oxidative stress parameters in infected mice. The infected non-treated group exhibited elevated serum levels of ALT, AST, total cholesterol, triglycerides, LDL, and MDA compared with the non-infected control group, reflecting hepatic injury, metabolic disturbance, and enhanced lipid peroxidation. For instance, Taha et al. (2023) reported that C. parvum infection in immunosuppressed mice caused a marked elevation in liver transaminases, lipid peroxidation, and antioxidant defense impairment. Similarly, Rahman et al. (2022) observed that cryptosporidiosis disrupted lipid metabolism and increased oxidative stress biomarkers in infected hosts, indicating that oxidative injury plays a central role in Cryptosporidium-induced pathogenesis. Conversely, treatment with S. arboricola extract, particularly at a higher dose (200 mg/kg), significantly reduced these altered biochemical parameters. The observed hepatoprotective and antioxidant effects can be attributed to the rich phytochemical composition of S. arboricola, notably its phenolic acids and flavonoids such as rosmarinic acid, quercetin, and kaempferol, which are well known for their free radical scavenging and lipid-stabilizing properties (Sharma et al., 2021; Pellati et al., 2004). These bioactive metabolites likely modulate antioxidant enzyme activity, suppress lipid peroxidation, and improve cellular redox balance (El-Feky and El Batanony, 2022).
Furthermore, the quantitative DNA fragmentation assay revealed that C. parvum infection induced marked DNA damage in the intestinal tissues of infected mice. This observation aligns with earlier reports indicating that Cryptosporidium infection can trigger apoptosis and DNA strand breaks through oxidative stress-mediated pathways and inflammatory responses (Atwa et al., 2020). In contrast, S. arboricola extract-treated mice exhibited a substantial reduction in DNA fragmentation. This improvement may be attributed to the strong antioxidant defense provided by S. arboricola phenolic constituents, which mitigate oxidative DNA damage by scavenging free radicals and stabilizing cellular membranes. Compounds such as rosmarinic acid, caffeic acid, and quercetin have been reported to attenuate DNA fragmentation by inhibiting ROS generation and modulating apoptotic signaling cascades (Ayna, 2021). Thus, the decline in DNA fragmentation following S. arboricola treatment indicates the genoprotective potential of the extract against Cryptosporidium-induced oxidative injury.
Based on the parasitological, biochemical, antioxidant, and histopathological findings, the anti-Cryptosporidium activity of S. arboricola leaf extract is likely mediated through multiple complementary mechanisms. Cryptosporidium parvum infection is associated with pronounced intestinal oxidative stress and inflammation, which contribute to epithelial damage and parasite persistence. The improvement in antioxidant parameters observed in treated groups suggests that phenolic acids and flavonoids in the extract may mitigate oxidative injury by scavenging reactive oxygen species and restoring redox balance (Bhagat et al., 2017). Flavonoids such as quercetin, kaempferol, naringenin, and hesperetin are known to suppress pro-inflammatory signaling pathways, including Nuclear Factor kappa-light-chain-enhancer of activated B cells and Cyclooxygenase-2, which may account for the marked histopathological recovery of intestinal tissues (Hämäläinen et al., 2007). Additionally, several of the identified phenolic compounds have been reported to exert direct antiprotozoal effects by interfering with parasite metabolism and viability, suggesting a possible direct inhibitory action against Cryptosporidium developmental stages (Soto-Sánchez, 2022). Collectively, the anti-Cryptosporidium efficacy of S. arboricola appears to arise from a synergistic interplay between antioxidant, anti-inflammatory, and direct antiparasitic effects.
In addition to its therapeutic efficacy, S. arboricola leaf extract demonstrated a favorable tolerability profile under the experimental conditions. The absence of elevations in serum ALT and AST levels in the treated groups, together with the normalization of lipid profile parameters and oxidative stress markers, indicates that the extract did not exert hepatotoxic or metabolic adverse effects. In contrast, treatment—particularly at the higher dose (200 mg/kg)—ameliorated the biochemical disturbances induced by C. parvum infection, suggesting a hepatoprotective rather than toxic effect. No abnormal clinical signs, behavioral changes, or mortality were observed in extract-treated mice throughout the experimental period. These findings are consistent with previous reports highlighting the relative safety of therapeutic doses of phenolic- and flavonoid-rich plant extracts and support the potential advantage of S. arboricola as a well-tolerated natural alternative or complementary agent in the management of cryptosporidiosis.
Although the present study demonstrated the promising anti-Cryptosporidium efficacy and favorable tolerability of S. arboricola leaf extract in an experimental mouse model, further investigations are required to fully confirm its therapeutic potential. Sub-chronic and chronic toxicity assessments should be included in future studies to establish long-term safety, as well as bioactivity-guided fractionation to identify the most active constituents.
Conclusion
The leaves of S. arboricola L. extract were found to contain high concentrations of numerous carbohydrates and pigments, such as chlorophyll, carotene, phenolics, and flavonoids. The extract demonstrated significant anti-C. parvum activity in experimentally infected mice, as evidenced by reduced oocyst shedding, improved histopathological architecture, and amelioration of oxidative stress and biochemical disturbances. Schefflera arboricola is a promising natural therapeutic candidate for the management of cryptosporidiosis.
Acknowledgments
The authors are grateful to their colleagues at the National Research Centre’s Pharmacognosy Department, Zoonotic Disease Department, and Parasitology and Animal Diseases Department. In addition, we thank our colleagues in the Faculty of Veterinary Medicine at Tripoli University in Libya for conducting this cooperative study.
Conflict of interest
The author declares no conflict of interest.
Funding
This research received no specific grant.
Authors’ contributions
Amal M. El-Feky performed the phytochemical analyses, interpreted the data, and prepared the original manuscript draft. Amany M. Mohamed, Nehal M. Khairy, and Raafat M. Shaapan conducted the biological experiments, evaluated the therapeutic efficacy, and analyzed and interpreted the data. Magdy A. A. Ayad as a corresponding author. All authors participated in the revision of the manuscript, approved the final version, and agreed to be accountable for all aspects of the work.
Data availability
The data supporting the findings of this study are available on request from the corresponding author, and no additional data sources are required.
References
Abd El Wahab, W.M., Shaapan, R.M., El-Naggar, E.M.B., Ahmed, M.M., Owis, A.I. and Ali, M.I. 2022. Anti-Cryptosporidium efficacy of Citrus sinensis peel extract: histopathological and ultrastructural experimental study. Exp. Parasitol. 243, 108412; doi:10.1016/j.exppara.2022.108412
Abouelsoued, D., Toaleb, N.I., Ibrahim, S., Shaapan, R.M. and Megeed, K.N.A. 2023. A Cryptosporidium parvum vaccine candidate effect on immunohistochemical profiling of CD4+, CD8+, Caspase-3 and NF-κB in mice. BMC. Vet. Res. 19(1), 216; doi: 10.1186/s12917-023-03699-w
Abouelsoued, D., Shaapan, R., Elkhateeb, R.M., Elnattat, W., Abd Elhameed, M., Hammam, A.M.M. and Hammam, A.M. 2020. Therapeutic efficacy of ginger (Zingiber officinale), ginseng (Panax ginseng) and sage (Salvia officinalis) against Cryptosporidium parvum in experimentally infected mice. Egypt. J. Vet. Sci. 51(2), 241–251; https://10.21608/EJVS.2020.24183.1152
Abu El Ezz, N.A., Khalil, F.A. and Shaapan, R.M. 2011. Therapeutic effect of onion (Allium cepa) and cinnamon (Cinnamomum zeylanicum) oils on cryptosporidiosis in experimentally infected mice. Glob. Vet. 7(2), 179–183; www.idosi.org/gv/GV7(2)11/13.pdf
Alday-Provencio, S., Diaz, G., Rascon, L., Quintero, J., Alday, E., Robles-Zepeda, R., Garibay-Escobar, A., Astiazaran, H., Hernandez, J. and Velazquez, C. 2015. Sonoran propolis and some of its chemical constituents inhibit in vitro growth of Giardia lamblia trophozoites. Planta. Med. 81(9), 742–747; doi:10.1055/s-0035-1545982
Atwa, M., Salem, N. and Mohamed, H. 2020. Effect of Cryptosporidium parvum infection on DNA damage and cytokines gene expression in blood of immunocompromised mice. Egypt. J. Zool. 73(73), 16–25; doi:10.21608/ejz.2020.24247.1021
Ayna, A. 2021. Caffeic acid prevents hydrogen peroxide-induced oxidative damage in SH-SY5Y cell line through mitigation of oxidative stress and apoptosis. BRATISL. MED. J. /. Bratisl. Lek. Listy. 122(2), 120; doi:10.4149/BLL_2021_018
Baba, S.A. and Malik, S.A. 2015. Determination of total phenolic and flavonoid content, antimicrobial and antioxidant activity of a root extract of Arisaema jacquemontii Blume. J. Taibah. Univ. Sci. 9(4), 449–454; doi:10.1016/j.jtusci.2014.11.001
Barakat, A.Z., Abdel-Aty, A.M., Salah, H.A., Ibrahim, M.K., Ibrahim, T.B., Shaapan, R.M. and Mohamed, S.A. 2024. Targeting Sarcocystis fusiformis cysteine protease as a possible approach to limit sarcocystosis infection. Egypt. J. Chem. 67(13), 911–924; doi:10.21608/ejchem.2024.325966.10587
Bhagat, M., Sood, S., Yadav, A., Verma, P., Manzoor, N., Chakraborty, D., Katoch, R. and Sangha, N. 2017. Alterations in oxidative stress parameters and its associated correlation with clinical disease on experimental Cryptosporidium parvum infection in Swiss albino mice. J. Parasit. Dis. 41(3), 707–712; doi:10.1007/s12639-016-0871-5
Bodnar, O.I., Viniarska, H.B., Vasilenko, O.V. and Grubinko, V.V. 2016. Pigment content of Chlorella vulgaris Beij. under influence of sodium selenite and metals ions. Biotechnol. Acta. 9(1), 71–78; doi:10.15407/biotech9.01.071
Caravedo, M.A. and White, A.C. 2023. Treatment of cryptosporidiosis: nitazoxanide yes, but we can do better. Expert. Rev. Anti-infect. Ther. 21(2), 167–173; doi:10.1080/14787210.2023.2160704
Chang, H.M. and Butt, P.P.H. 1986. Pharmacology and Applications of Chinese Materia Medica. World Scientific Press, Singapore, Singapore: World Scientific Press, Vol. I, p: 304.
Deepa, H.R. and NALINI, M.S. 2013. Phytochemical screening, total phenolic content and in vitro antioxidant studies of leaf, bark and flower extracts of Schefflera spp. (Araliaceae). J. App. Pharm. Sci. 3(11), 94–98; doi:10.7324/JAPS.2013.31117
Dubois, M., Gilles, K.A., Hamilton, J.K., Rebers, P.A. and Smith, F. 1956. Colorimetric method for determination of sugars and related substances. Anal. Chem. 28, 350–356; doi:10.1021/ac60111a017
Elfadaly, H.A., Hassanain, N.A., Hassanain, M.A., Barakat, A.M. and Shaapan, R.M. 2018. Evaluation of primitive ground water supplies as a risk factor for the development of major waterborne zoonosis in Egyptian children living in rural areas. J. Infect. Public. Health. 11(2), 203–208; doi:10.1016/j.jiph.2017.07.025
El-Feky, A. and El Batanony, M. 2022. Potential phytoconstituents of some fruit and vegetable peels against oxidative damage, inflammatory and cytotoxic diseases. Egypt. J. Chem. 65(3), 261–272; doi:10.21608/ejchem.2021.82086.4047
El-Feky, A.M., Aboulthana, W.M., El-Sayed, A.B. and Ibrahim, N.E. 2017. Chemical and therapeutic study of Nannochloropsis oculata on spleen of streptozotocin induced diabetes in rats. Der. Pharma. Chem. 9, 36–43. Available via http://www.derpharmachemica.com/archive.html
El-Hagrassi, A.M., Osman, A.F., El-Naggar, M.E., Mowaad, N.A., Khalil, S. and Hamed, M.A. 2022. Phytochemical constituents and protective efficacy of Schefflera arboricola L. leaves extract against thioacetamide-induced hepatic encephalopathy in rats. Biomark 27(4), 375–394; doi:10.1080/1354750x.2022.2048892
Fabre, N., Rustan, I., De Hoffmann, E. and Quetin-Leclercq, J. 2001. Determination of flavone, flavonol, and flavanone aglycones by negative ion liquid chromatography electrospray ion trap mass spectrometry. J. Am. Soc. Mass Spectrom. 12, 707–715; doi:10.1016/S1044-0305(01)00226-4
Farag, M.A., Rasheed, D.M., Kropf, M. and Heiss, A.G. 2016. Metabolite profiling in Trigonella seeds via UPLC-MS and GC-MS analyzed using multivariate data analyses. Anal. Bioanal. Chem. 408, 865–878; doi:10.1007/s00216-016-9910-4
Gardana, C., Scaglianti, M., Pietta, P. and Simonetti, P. 2007. Analysis of the polyphenolic fraction of propolis from different sources by liquid chromatography-tandem mass spectrometry. J. Pharm. Biomed. Anal. 45, 390–399; doi:10.1016/j.jpba.2007.06.022
Ghazy, A.A., Abdel-Shafy, S. and Shaapan, R.M. 2015b. Cryptosporidiosis in animals and man: 2. Diagnosis. As. J. Epidemiol. 8(4), 84–103; doi:10.3923/aje.2015.84.103
Ghazy, A.A. 2015b. Cryptosporidiosis in animals and man: 2. Diagnosis. As. J. Epidemiol. 8(4), 84–103; doi:10.3923/aje.2015.84.103
Ghazy, A.A., Abdel-Shafy, S. and Shaapan, R.M. 2016. Cryptosporidiosis in Animals and Man: 3. Prevention and control. As. J. Epidemiol. 9(1-3), 1-9; doi:10.3923/aje.2016.1.9
Guo, F.J., Chen, P., Peng, S.Y. and Li, Y.C. 2006. Triterpenoid Saponins from Schefflera arboricola. Helvetica Chim. Acta 89(3), 468–474; doi:10.1002/hlca.200690047
Hämäläinen, M., Nieminen, R., Vuorela, P., Heinonen, M. and Moilanen, E. 2007. Anti‐inflammatory effects of flavonoids: Genistein, kaempferol, quercetin, and daidzein inhibit STAT‐1 and NF‐κB activations, whereas flavone, isorhamnetin, naringenin, and pelargonidin inhibit only NF‐κB activation along with their inhibitory effect on iNOS expression and NO production in activated macrophages. Mediat. Inflamm. 2007(1), 045673; doi: 10.1155/2007/45673
Hassanain, M.A., Khalil, F.A., Abdel-Razik, K.A. and Shaapan, R.M., 2011. Prevalence and molecular discrimination of Cryptosporidium parvum in calves in Behira Provinces, Egypt. Res. J. Parasitol. 6 (3), 101–108; doi: 2011.101.108
Hossain, M.B., Rai, D.K., Brunton, N.P., Martin-Diana, A.B. and Barry-Ryan, C. 2010. Characterization of phenolic composition in Lamiaceae spices by LC–ESI-MS/MS. J. Agric. Food. Chem. 58, 10576–10581; doi:10.1021/jf102042g
Innes, E.A., Chalmers, R.M., Wells, B. and Pawlowic, M.C. 2020. A one-health approach to tackle cryptosporidiosis. Trends Parasitol. 36(3), 290–303; doi:10.1016/j.pt.2019.12.016
Javed, K. and Alkheraije, K.A. 2023. Cryptosporidiosis: a foodborne zoonotic disease of farm animals and humans. Pak. Vet. J. 43(2), 213–223. Available via http://pvj.com.pk/pdf-files/23-092.pdf
Kečkeš, S., Gašić, U., Veličković, T.C., Milojković-Opsenica, D., Natić, M. and Tešić, Z. 2013. The determination of phenolic profiles of Serbian unifloral honeys using ultra-high-performance liquid chromatography/high resolution accurate mass spectrometry. Food. Chem. 138, 32–40; doi:10.1016/j.foodchem.2012.10.025
Kyriakopoulou, E., Tsakni, A., Korakidis, E., Mpekoulis, G., Kalliampakou, K.I., Polanska, M., Van Impe, J.F.M., Tsakali, E., Houhoula, D. and Vassilaki, N. 2025. Evaluation of polyphenolic compounds common in Greek medicinal plants for their antioxidant effects and antiviral activity against dengue and yellow fever viruses. Antioxidants 14, 1103; doi:10.3390/antiox14091103
Liu, X., Liu, D.F. and Liu, H.B. 2011. Screening of anti-inflammatory and analgesic extraction from Schefflera arboricola Hayata. Anhui Med. Pharma. J. 12, 10–16.
Loannou, Y.A. and Chen, F.W. 1996. Quantitation of DNA fragmentation in apoptosis. Nucleic. Acids. Res. 24(5), 992–993; doi:10.1093/nar/24.5.992
Lombardelli, J.A., Tomazic, M.L., Schnittger, L. and Tiranti, K.I. 2019. Prevalence of Cryptosporidium parvum in dairy calves and GP60 subtyping of diarrheic calves in central Argentina. Parasitol. Res. 118(7), 2079–2086; doi:10.1007/s00436-019-06366-y
Lütjohann, D., Klör, H.U. and Stellaard, F. 2023. Measurement of serum low density lipoprotein cholesterol and triglyceride-rich remnant cholesterol as independent predictors of atherosclerotic cardiovascular disease: possibilities and limitations. Nutrients 15(9), 2202; doi:10.3390/nu15092202
Melek, F.R., Miyase, T., Abdel Khalik, S.A. and El-Gindi, M.R. 2003. Triterpenoid saponins from Schefflera arboricola. Phytochem. 63(4), 401–407; doi: 10.1016/s0031-9422(03)00117-1
Mead, J.R. and Mcnair, N. 2006. Antiparasitic activity of flavonoids and isoflavones against Cryptosporidium parvum and Encephalitozoon intestinalis. FEMS. Microbiol. Lett. 259(1), 153–157; doi:10.1111/j.1574-6968.2006.00263.x
Oliveira, B.C.M., Bresciani, K.D.S. and Widmer, G. 2019. Deprivation of dietary fiber enhances susceptibility of mice to cryptosporidiosis. PLos Negl. Trop. Dis. 13(9), e0007411; doi:10.1371/journal.pntd.0007411
Pellati, F., Benvenuti, S., Magro, L., Melegari, M. and Soragni, F. 2004. Analysis of phenolic compounds and radical scavenging activity of Echinacea spp. J. Pharm. Biomed. Anal. 35(2), 289–301; doi:10.1016/S0731-7085(03)00645-9
Pellati, F., Orlandini, G., Pinetti, D. and Benvenuti, S. 2011. HPLC-DAD and HPLC–ESI-MS/MS methods for metabolite profiling of propolis extracts. J. Pharm. Biomed. Anal. 55, 934–948; doi:10.1016/j.jpba.2011.03.024
Plunkett, G.M., Lowry Ii, P.P., Frodin, D.G. and Wen, J. 2005. Phylogeny and geography of Schefflera: pervasive polyphyly in the largest genus of Araliaceae. Ann. Missouri. Bot. Gard. 92, 202–224. Available via https://www.jstor.org/stable/3298514
Rahman, S.U., Zhou, K., Zhou, S., Sun, T., Mi, R., Huang, Y., Han, X., Gong, H. and Chen, Z. 2022. Curcumin mitigates Cryptosporidium parvum infection through modulation of gut microbiota and innate immune-related genes in immunosuppressed neonatal mice. Microb. Pathog. 164, 105424; doi:10.1016/j.micpath.2022.105424
Ristivojević, P., Trifković, J., Gašić, U., Andrić, F., Nedić, N., Tešić, Z. and Milojković‐Opsenica, D. 2015. Ultrahigh‐performance liquid chromatography and mass spectrometry (UHPLC–LTQ/Orbitrap/MS/MS) study of phenolic profile of Serbian poplar type propolis. Phytochem. Anal. 26(2), 127–136; doi:10.1002/pca.2544
Ruan, J., Yan, J., Zheng, D., Sun, F., Wang, J., Han, L., Zhang, Y. and Wang, T. 2019. Comprehensive chemical profiling in the ethanol extract of Pluchea Indica aerial parts by liquid chromatography/mass spectrometry analysis of its silica gel column chromatography fractions. Molecules 24(15), 278; doi:10.3390/molecules24152784
Saad-Alla, G.I., Shaapan, R.M., Varjabedian, K.G., Abdel-Ghaffar, F.A. and Khalil, F.A.M. 2022. Cryptosporidiosis in Clarias gariepinus fish: incidence and molecular aspect. Egypt. J. Aquatic Biol. Fish. 26(3), 775–786; doi: 10.21608/ejabf.2022.246473
Shaapan, R., Elfadaly, H., Elaadli, H., Salman, M.B., Maher, A. and Barakat, A. 2023. Re-shedding trials of Toxoplasma gondii oocysts from experimentally infected kittens reference to strain types I, II and III; zoonotic and histopathological confirm. J. Adv. Vet. Res. 13(9), 1776–1781; https://advetresearch.com/index.php/AVR/article/view/1543
Shaapan, R.M., Abdel-Ghaffar, F.A., Varjabedian, K.G. and Saad-Alla, G.I. 2022. Prevalence and molecular epidemiology of Cryptosporidium infection in Clarias gariepinus fish in Egypt. Acta Parasitologica 67(1), 437–445; doi:10.1007/S11686-021-00483-4
Shaapan, R.M., Hassanain, M.A., A.m. Khali, F. and Abdel-Razi, K.A. 2011. Prevalence and molecular discrimination of Cryptosporidium parvum in calves in Behira Provinces, Egypt. Res. J. Parasitol. 6(3), 101–108; http://doi=jp.2011.101.108
Sharma, N., Biswas, S., Al-Dayan, N., Alhegaili, A.S. and Sarwat, M. 2021. Antioxidant role of kaempferol in prevention of hepatocellular carcinoma. Antioxidants 10(9), 1419; doi:10.3390/antiox10091419
Shoukry, N., Mekky, E.A., Salah, I.M., Darwish, A.B. and Aly, I. 2025. Immunological alterations of Lactoferrin, Silver Nanoparticles, and Nitazoxanide therapy in Immunocompromised Murine Cryptosporidiosis. Front. Sci. Res. Technol. 12, 1–6; doi:10.21608/fsrt.2025.393245.1169
Soto-Sánchez. and J. 2022. Bioactivity of natural polyphenols as antiparasitic agents and their biochemical targets. Mini-Rev. Med. Chem. 22(20), 2661–2677; doi:10.2174/1389557522666220404090429
Taha, N.M., Zalat, R.S., Khaled, E. and Elmansory, B.M. 2023. Evaluation of the therapeutic efficacy of some essential oils in experimentally immunosuppressed mice infected with Cryptosporidium parvum. J. Parasit. Dis. 47(4), 733–743; doi:10.1007/s12639-023-01621-7
Toaleb, N. and Shaapan, R. 2024. Zoonotic Protozoan parasites infecting camels, diagnosis and control–a review. Egypt. J. Vet. Sci. 55(4), 1131–1142; doi:10.21608/ejvs.2023.251609.1686
Wang, Y., Khan, F.A., Siddiqui, M., Aamer, M., Lu, C. and Choudhary, M.I. 2021. The genus Schefflera: a review of traditional uses, phytochemistry and pharmacology. J. Ethnopharmacol. 279, 113675; doi:10.1016/j.jep.2020.113675
Wang, Y., Liang, D., Khan, F.A., Zhang, C.L., Liu, Y.F., Chen, R.Y., Choudhary, M.I. and Yu, D.Q. 2020. Chemical constituents from Schefflera leucantha R.vig. (Araliaceae). Biochem. Syst. Ecol. 91, 104076; doi: 10.1016/j.bse.2020.104076
Xiao, L., Fayer, R., Ryan, U. and Upton, S.J. 2004. Cryptosporidium taxonomy: recent advances and implications for public health. Clin. Microbiol. Rev. 17(1), 72–97; doi:10.1128/cmr.17.1.72-97.2004
Zeng, Y., Li, S., Wang, X., Gong, T., Sun, X. and Zhang, Z. 2015. Validated LC-MS/MS method for the determination of Scopoletin in rat plasma and its application to pharmacokinetic studies. Molecules 20, 18988–19001; doi:10.3390/molecules201018988
Zhao, Z., Matsunami, K., Otsuka, H., Shinzato, T., Takeda, Y., Kawahata, M. and Yamaguchi, K. 2010. Schefflerins A—G, New Triterpene Glucosides from the Leaves of Schefflera arboricola. Chem. Pharm. Bull. 58(10), 1343–1348; doi:10.1248/cpb.58.1343






