




| Research Article | ||
Open Vet. J.. 2026; 16(5): 2970-2978 Open Veterinary Journal, (2026), Vol. 16(5): 2970-2978 Research Article Stage-specific gene expression profiling of Toxocara canis in canine and human hosts: Implications for the pathogenesis and transmission of zoonotic diseasesAmal Hassan Abd Al_Shabbani1, Saafa Ressan Abdullah Al-Kaeebi2, Miras Hasan Madhloom3 and Monyer Abdulamier Abd Alfatlawi4*1Clinical Laboratory Sciences, College of Pharmacy, University of Al-Qadisiyah, Al Diwaniyah, Iraq 2Nursing College, University of Al-Qadisiyah, Al Diwaniyah, Iraq 3College of Science, University of Al-Qadisiyah, Al Diwaniyah, Iraq 4Department of Veterinary Microbiology, College of Veterinary Medicine, University of Al-Qadisiyah, Al Diwaniyah, Iraq *Corresponding Author: Monyer Abdulamier Abd Alfatlawi. Department of Veterinary Microbiology, College of Veterinary Medicine, University of Al-Qadisiyah, Al Diwaniyah, Iraq. Email: monyerr.abd [at] qu.edu.iq Submitted: 17/11/2025 Revised: 14/03/2026 Accepted: 24/03/2026 Published: 31/05/2026 © 2025 Open Veterinary Journal
ABSTRACTBackground: Toxocara canis is a zoonotic nematode that mainly infects dogs but can produce visceral, ocular, and neurological toxocariasis in humans. The parasite completes its life cycle within dogs. The molecular processes that enable T. canis to adapt to its host remain largely unknown despite the significance of the parasite to public health. Aim: To understand the molecular basis of immune evasion, tissue migration, and survival strategies, researchers analyzed T. canis gene expression during different life stages within canine and human hosts. Methods: Third-stage larvae (L3) and adult worms were collected from infected tissues at intervals of 7, 14, 28, and 35 days post-infection using both experimentally infected dogs (definitive hosts) and murine models (human equivalent accidental hosts). RNA was extracted, and quantitative real-time PCR was performed to measure the expression of three key virulence genes: Toxocara excretory-secretory protein 1 (TcES-1), Toxocara cysteine protease-like gene (TcCPL-1), and Toxocara heat shock protein 90 (TcHSP-90) were examined for their roles in worm virulence. Results: Human-model larvae showed significantly higher levels of TcES-1 and TcCPL-1 expression than canine larvae, indicating stronger immune modulation and tissue invasion abilities in incidental hosts with statistical significance (p < 0.001). Human-model larvae displayed significantly higher levels of TcHSP-90 expression than both dog-derived larvae and adult worms, indicating stress-response adaptation in non-permissive hosts. Conclusion: The findings of this study reveal how T. canis adapts to different hosts at the molecular level and identify new opportunities for the development of diagnostic and therapeutic approaches to manage zoonotic toxocariasis. Keywords: Cysteine protease, Excretory-secretory proteins, Host adaptation, Toxocara canis, Zoonosis. IntroductionToxocara canis is an important parasitic nematode of canids with a worldwide distribution. It is recognized as a common gastrointestinal nematode of domestic dogs, representing a key pathogen that affects canine health. Toxocara canis is also a zoonotic pathogen with significant public health implications. In human hosts, the parasite can lead to a range of clinical consequences (toxocariasis), such as visceral larva migrans (VLMs) syndrome, ocular larva migrans, and neurotoxocariasis (Zhu et al., 2015; Schwartz et al., 2022). People are unintentionally infected with T. canis by oral ingestion of the infective eggs present in the environment through contact with contaminated soil. The widespread presence of T. canis larvae in tissues of diverse animal hosts can threaten the lives of diverse hosts and lead to the pathological consequences mentioned above in new and accidental hosts, including humans (Dar et al., 2016). Additionally, the contaminated environments by T. canis eggs and the risks of accidental ingestion in humans via infected dogs is of concern and is emphasized as a One Health subject. Therefore, there is a need for efficient control, and people play a key role in this regard through education and understanding of parasite epidemiology, pathogenesis, and disease prevention (Tylkowska et al., 2024). Toxocara canis, one of the most common and prevalent parasitic nematodes infecting domestic dogs globally, has attracted medical and veterinary researchers for many years (Saichenko et al., 2021). The genus Toxocara comprises the most frequent zoonotic nematode helminths, T. canis and Toxocara cati, which infect various intermediate or paratenic hosts. This dog roundworm is one of the most common zoonotic helminthes, infecting dogs worldwide three. Furthermore, the role of free-ranging dogs, representing a very serious risk of infection of the human population with this zoonosis, as well as the persistence of its eggs in various ecosystems, has been discussed (Jahanmahin and Borji, 2023). Toxocara canis infection is mainly associated with a high proportion of juvenile and young adult dogs, particularly during the second month of life; patent infections are less common (up to 37%) in adult pets. The prevalence and intensity of T. canis are generally associated with the environmental conditions present in a widely different geographic area. Feeding dogs with noncommercial food or leftovers significantly increases the risk of T. canis infection. However, the effect is indirect and not direct, probably associated with the worse health and body condition of dogs fed in this way. Globally, the number of dogs and human infection rates show a similar pattern; this is especially noticeable in the second half of the age range (Holland, 2023). Currently, mass dog deworming programs using macrocyclic lactones are conducted in several countries. Notwithstanding these measures, a high prevalence of T. canis infection is still observed in the youngest dogs. Among the important biological and environmental factors that intrinsically influence the circulation of T. canis is the persistence of their eggs in the long-term environment. Sporulation of live eggs is essential for maturation. The feces of T. canis-infected dogs excrete eggs that are not sporulated. Under suitable conditions, ripe/ mature eggs sporulate in 10–14 days and become infectious (embryonated eggs) for the next host, whereas unsuitable eggs may remain in the environment for months, over 1 year, and even for 2 years (Dar et al., 2016; Saelens et al., 2022). Zoonotic transmission of T. canis is a complex issue involving interactions among the environment, pathogen, companion host, and human host. Dogs are commonly infected with the parasites, and their eggs are broadly shed, particularly in the feces. Toxocara canis infection-related illness in humans has been well documented, particularly in the context of children, as they tend to be more frequently infected with the parasites (Schwartz et al., 2021; Airs et al., 2023). The increasing recognition of the general importance of companion animals in sustaining human health adds to the ratification of the One Health approach (Gelmi-Candusso et al., 2024). Understanding the health impacts of T. canis infections on dogs is of greater importance because of the potential for zoonotic transmission. Zoonotic transmission to humans may cause toxocariasis, a neglected and emergent parasitic disease (Henke et al., 2023; Jahanmahin and Borji, 2023). Upon the accidental ingestion of embryonated eggs from the environment, the larvae will hatch, penetrate the intestinal wall, or remain entrapped and migrate to various body tissues through the blood or the lymphatic system (Qualizza et al., 2011). In human hosts, Toxocara spp. larvae do not develop into adults, their movement is disordered, and tissue and organ damage are induced (Magnaval et al., 2022). The most common symptoms, which can occur almost under every clinical condition, are fever or fever-like symptoms, malaise, headaches, and cutaneous eruptions (Cabral Monica et al., 2022). Materials and MethodsAnimalsSix healthy and parasite-free puppies, aged 8–10 weeks, were used as definitive hosts and randomly assigned to the infected (n=4) and control (n=2) groups. In addition, 30 BALB/c mice (both males and females, age 6–8 weeks) were used as accidental hosts, randomly assigned to the infected (n=24) and control (n=6) groups. Sample collectionFresh fecal samples were obtained from naturally infected dogs that tested positive for T. canis eggs via microscopic examination and fecal flotation. The isolation of eggs was performed by suspending fecal material in saturated NaCl solution and then centrifuging the mixture at 1,500 × g for 5 minutes to concentrate the eggs. The supernatant was transferred into a Baermann funnel to enable egg sedimentation before washing the eggs multiple times with sterile distilled water. Under controlled conditions, embryonation occurred when purified eggs were incubated for 4 weeks at 27°C in 0.5% formalin solution. The third-stage larvae (L3) containing embryonated eggs were stored at 4°C before being used for experimental infections. For reliable infection of all subjects, we characterized the quantified egg suspension homogenate of the sampled feces. The measured egg suspension was obtained using the Neubauer counting chamber with the light microscope at 400× magnification. The egg counts were homogenized, measured, and repeated thrice independently. The average egg count was measured, resulting in an average of 12,450 ± 1,320 eggs per gram of feces from natural infection samples. After embryonation, the egg viability was microscopically evaluated, and 82% of the assessed eggs were L3 larvae. The final larval doses were set to these values (1,000 and 500 eggs for dogs and 500 eggs for mice). Experimental infection in canine and murine modelsFor infection experiments, researchers investigated host-specific adaptation using dogs as definitive hosts and murine models as accidental human-equivalent hosts. Animals were maintained under pathogen-free conditions following the ethical guidelines for animal research. Researchers administered 1,000 embryonated eggs to dogs through food and 500 larvae to mice using gastric gavage with a soft needle. The murine model was selected to replicate human visceral toxocariasis because T. canis larvae migrate through tissues for extended periods in accidental hosts. The control groups consisted of non-infected dogs and mice that received sterile saline and infected animals that received eggs. Recovery of larval and adult worms from host tissuesDuring the gene expression analysis of different parasite stages, samples were collected at four distinct time intervals after infection. The researchers performed euthanasia on infected mice at 7, 14, and 28 days post-infection (dpi) through CO₂ asphyxiation and cervical dislocation before extracting organs with a stereomicroscope. Tissue samples from the liver, lungs, and brain were digested with a pepsin-HCl solution (pH 2.0) at 37°C for 4 hours to isolate L3 larvae, which were then separated through sedimentation and cleaned with phosphate-buffered saline (PBS) (pH 7.4). Adult worms were extracted from dogs at 35 dpi by performing necropsies and isolating parasites from the small intestine contents. The collected larvae and adult worms were washed three times with sterile PBS to eliminate host tissue debris before RNA extraction. RNA extractionThe TRIzol reagent from Thermo Fisher Scientific was used according to the manufacturer’s instructions to extract total RNA from both L3 larvae derived from mice and adult worms collected from dogs. The research team homogenized samples containing 1 × 10⁶ larvae or 100 adult worms with 1 ml of TRIzol reagent using a Bead Ruptor Homogenizer from Omni International. After adding chloroform (200 µl), the mixture was centrifuged at 12,000 × g for 15 minutes at 4°C. The aqueous phase was transferred to a new tube before adding 500 µl of isopropanol to precipitate RNA, which was then incubated at −20°C for 30 minutes, followed by centrifugation at 12,000 × g for 10 minutes. Researchers washed the RNA pellet twice using 75% ethanol and then air-dried it before resuspending it in RNase-free water. The Thermo Fisher Scientific’s NanoDrop 2000 Spectrophotometer was used to measure RNA purity and concentration by achieving an A260/A280 ratio between 1.8 and 2.2. The integrity of the RNA was confirmed through 1.5% agarose gel electrophoresis, which showed intact ribosomal RNA bands. cDNA synthesis and reverse transcriptionThe RevertAid First Strand cDNA Synthesis Kit from Thermo Fisher Scientific was used to synthesize cDNA from high-quality RNA. The synthesis reaction had 1 µg of total RNA with random hexamer primers, 0.5 mM dNTPs, 200 U of reverse transcriptase, and 20 U of RNase inhibitor in a total 20 µl volume. The reaction was incubated at 42°C for 1 hour before enzyme inactivation at 70°C for 5 minutes. The cDNA sample remained frozen at −20°C until its next application. Quantitative real-time PCR (qRT-PCR) is the standard approach for analyzing gene expression levels. A QuantStudio 5 qRT-PCR System (Applied Biosystems) was used to perform qRT–PCR analysis of gene expression levels for three primary virulence-associated genes. The genes selected were: Toxocara excretory-secretory protein 1 (TcES-1): This protein operates as an immune modulator by suppressing host cytokine activity. Cysteine protease-like gene (TcCPL-1): The TcCPL-1 gene plays a role in breaking down host proteins and penetrating tissue barriers. Heat shock protein 90 (TcHSP-90): Facilitates parasite survival under host-induced stress conditions. Researchers employed glyceraldehyde-3-phosphate dehydrogenase (TcGAPDH) as an internal control to normalize gene expression. The primer sequences were designed using NCBI Primer-BLAST and were confirmed to match specific targets. Primer sequences are as follows: TcES-1 forward:Sequences are TcES-1 Forward: 5’-AGCTGACGGTGAATCGTACG-3’ and Reverse: 5’-TCCACGTTGAGCTTTGGAAC-3’. TcCPL-1 5’-CGCGGATCCATGCGCGCGGCAATGC-3 and 5’-CCGGAATTCTTAAGAGGTCTCCTTGATTTC The qPCR reactions were conducted in 20 µl volumes with components including 10 µl SYBR Green Master Mix and 1 µl cDNA (corresponding to 50 ng RNA input). The reaction also contained 0.5 µl of each primer at 10 µM concentration and 8 µl of nuclease-free water. The thermal cycling conditions were as follows: initial denaturation at 95°C for 10 minutes, followed by 40 cycles of amplification: 95°C for 15 seconds (denaturation), 60°C for 30 seconds (annealing), and 72°C for 30 seconds (extension). The melting curve analysis between 60°C and 95°C confirmed amplicon specificity. The analysis of each sample included three replicates and controls without any template material. Statistical and bioinformatics analysisRelative gene expression was determined by applying the 2^(-ΔΔCt) method and using TcGAPDH as the normalization reference. Statistical significance was determined using one-way ANOVA together with Tukey’s multiple comparisons test, with the significance cutoff set at p < 0.05. The creation of data visualizations used GraphPad Prism 9, which produced: The study used bar plots to visualize comparisons of expression levels between various host-parasite conditions. Violin plots displaying gene expression variation across different biological replicates. Heatmaps were used to demonstrate fold-change variations between different parasite stages. Ethical approvalThe Institutional Animal Care and Use Committee of the College of Veterinary Medicine, University of Al-Qadisiyah approved the experimental protocol (Approval No.: VET-ACUC-2023). The protocol conformed to the national and institutional guidelines for laboratory animal use. ResultsAt specific time points during the infections, larvae recovered from murine hosts were distributed in the livers, lungs, and brains. Adult worms were recovered from the small bowel of the canine hosts at 35 dpi. As such, gene expression analysis pertains to larvae pooled from tissues other than the intestine of murine hosts and adult worms from the intestine of the definitive canine hosts. Toxocara canis virulence gene levelsExpression analysis of T. canis virulence genes revealed significant differences between larvae collected from canine definitive hosts and accidental human-equivalent murine hosts. The study monitored gene expression patterns in larvae at 7, 14, and 28 dpi, along with adult worms collected from dogs at 35 dpi. Researchers determined the relative gene expression levels of TcES-1, TcCPL-1, and TcHSP-90 through qRT–PCR analysis and normalization against the housekeeping gene TcGAPDH followed by analysis via the 2^(-ΔΔCt) method. The results revealed that transcriptional regulation depends on the host, as murine larvae showed significantly different expression profiles than canine larvae (Fig. 1A and B).
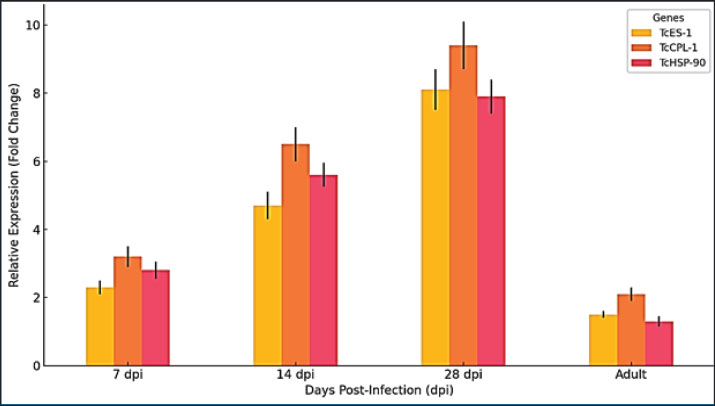
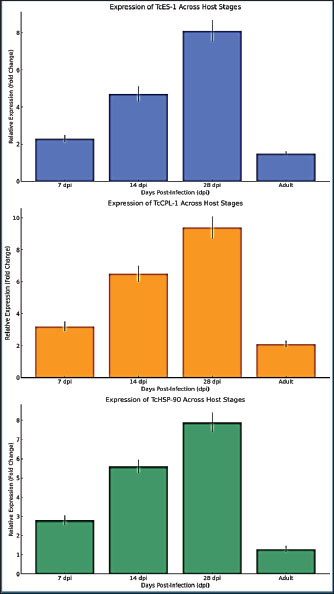
Fig. 1. Expression levels of TcES-1 (A) and TcCPL-1 (B) in larvae recovered from T. canis infected murine accidental hosts and from canine definitive hosts at 7-, 14-, and 28- dpi and in adult worms from dogs at 35 dpi. Expression values were normalized to TcGAPDH and determined by the 2^(-ΔΔCt) method. Data represent the mean ± SD of three biological replicates. Expression values were statistically analyzed by one-way analysis of variance and Tukey’s post hoc test, with levels of significance indicated by *p < 0.05, **p < 0.01, and ***p < 0.001. Enhanced TcES-1 protein expression in murine host larvae indicates increased immune modulation activityThe TcES-1 gene, which is responsible for producing an immune-suppressing excretory-secretory (ES) protein, was expressed at higher levels in larvae from murine hosts than in larvae from canine hosts. The gene expression level in murine-derived larvae reached 2.3 times that of canine-derived larvae at 7 dpi (p < 0.05). The expression levels diverged more significantly at 14 dpi, reaching a 4.7-fold increase (p < 0.01), and reached peak expression at 28 dpi with an 8.1-fold increase (p < 0.001). TcES-1 expression in canine-derived larvae remained stable and showed only a minor 1.5-fold growth between 7 and 28 dpi, which was not statistically significant (p=0.09) (Fig. 1A and B). TcES-1 expression continues to increase within murine larvaeThis indicates that the parasite produces immunomodulatory proteins in non-permissive hosts to escape host immune defenses. Toxocara canis larvae survive longer in murine tissues because they produce more immune-suppressive molecules compared with their fast intestinal development in dogs, which helps them escape host defenses. TcES-1 is a crucial element for the persistence of chronic toxocariasis in human hosts, where larvae survive for extended durations (Fig. 1A and B). TcCPL-1 expression in murine hosts shows significant elevation alongside improved tissue migration capabilityMurine-derived larvae displayed higher expression of the TcCPL-1 gene, which encodes the cysteine protease-like protein essential for tissue penetration and host protein breakdown, than canine-derived larvae or adult worms. At 7 dpi, the expression level of TcCPL-1 reached 3.2 times its baseline in murine-derived larvae, which was statistically significant (p < 0.01) when compared to canine-derived larvae. The gene expression reached a 6.5-fold increase by day 14 dpi, indicating ongoing protease activity as larvae moved through host tissues. The expression of TcCPL-1 remained high at 28 dpi with a 9.4-fold increase (p < 0.001), indicating that larvae need ongoing proteolytic activity to break down extracellular matrix proteins and move through host tissues. Dog-derived larvae exhibited consistently lower TcCPL-1 expression, which reached a maximum 2.1-fold increase at 14 dpi but was statistically insignificant (p=0.07) before decreasing. When worms reached adulthood by 35 dpi, they showed minimal TcCPL-1 expression because proteolytic activity becomes unnecessary for tissue invasion once larvae reach their intestinal stage within the definitive host. The results demonstrate that TcCPL-1 functions as a key factor for larval migration in accidental hosts, which aligns with the existing findings that T. canis larvae remain active and migrate throughout human tissues, leading to chronic VLM. The murine host environment triggers stress response adaptations in parasites, as shown by the host-specific expression pattern of TcHSP-90Murine-derived larvae displayed significant upregulation of the TcHSP-90 gene encoding heat shock proteins for parasite survival under host-induced stress compared with both canine-derived larvae and adult worms. Murine larvae exhibited a 2.8-fold elevation in TcHSP-90 levels 7 dpi, demonstrating an early stress response to their accidental host surroundings. The gene expression reached 5.6 times its original level by 14 dpi (p < 0.01) and further increased to 7.9 times its original level by 28 dpi (p < 0.001) (Fig. 2A).
Fig. 2. Expression levels of TcHSP-90 in murine and canine larvae at various time points during infection Box plots show the median and interquartile range and identify outliers. One-way analysis of variance followed by Tukey’s multiple comparison test was used to conduct statistical analyses. TcHSP-90 expression in canine larvae demonstrated minimal variation, which included a 1.3-fold increase at 7 dpi without statistical significance (p=0.12) and returned to baseline by 14 dpi. The near-absence of TcHSP-90 expression in adult worms implies that heat shock protein activity functions mainly to help organisms survive hostile environments instead of supporting adult worms in the definitive host’s gut (Fig. 2B). The results demonstrate the crucial function of TcHSP-90 in stress adaptation, which helps T. canis larvae survive oxidative stress and inflammatory responses as well as temperature changes when they reside in non-permissive hosts. Murine-derived larvae showed significantly elevated expression levels, indicating that human infections cause increased physiological stress to larvae, leading to a requirement for greater chaperone function to preserve protein stability and cellular homeostasis (Fig. 2C). Statistical evaluation demonstrates significant host-dependent variations in gene expression patternsStatistical analysis using one-way analysis of variance and Tukey’s multiple comparisons test established that differences in the expression of all three genes among the experimental groups reached statistical significance. TcES-1: F=18.92, p < 0.001, TcCPL-1: F=23.51, p < 0.001, and TcHSP-90: F=15.76, p < 0.001. Murine-derived larvae at 28 dpi displayed peak expression levels of three genes, which differed significantly from those of canine-derived larvae (p < 0.001) and adult worms (p < 0.001) according to post-hoc analysis. The data showed consistent differences across biological replicates since the average coefficient of variation remained under 7%, demonstrating highly reproducible results (Fig. 3).
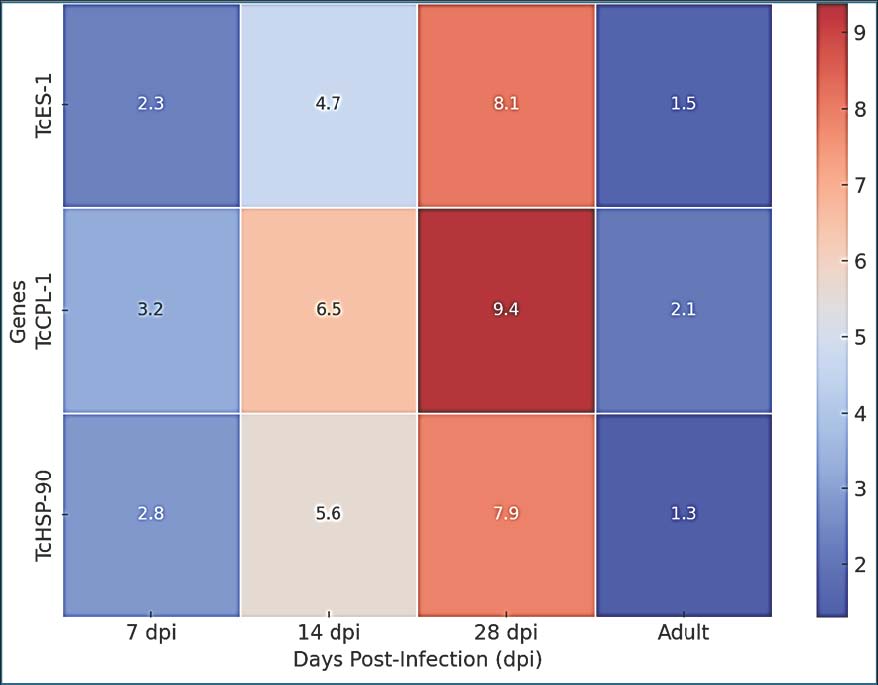
Fig. 3. Heatmap of fold change for TcES-1, TcCPL-1, and TcHSP-90 expression across different combinations of host stages. The degree of color saturation reflects transcriptional upregulation compared with the baseline expression in canine larvae. DiscussionThe gene expression analysis data illustrated in Fig. 1 compared transcription levels in both murine accidental and canine definitive hosts at different stages of infection. The expression of genes in hosts at different stages of infection is due to the specific host and not the infection stage. Toxocara canis is a major zoonotic parasite that infects both human hosts and animals. It causes multiple pathological outcomes through larval migration, resulting in visceral toxocariasis, ocular toxocariasis, and neurological toxocariasis. Transcriptomic and molecular research has uncovered essential information about gene expression mechanisms that help the parasite survive while adapting to hosts and evading their immune systems. The transcriptomic analysis of spleen and liver tissues in Beagle dogs demonstrated that T. canis infection triggers changes in the expression of lncRNA and mRNA, which impact both immune responses and metabolic pathways (Xu et al., 2022; Zou et al., 2023). The results of this research show that murine-derived larvae demonstrate an improved capability to both alter host immune functions and navigate tissue environments through the increased expression of TcES-1 and TcCPL-1 genes compared to canine hosts. According to observed differential expression patterns across different host species, genetic expression modulation in T. canis appears to depend on host-specific environmental factors. Studies on neurotoxocarosis reveal extensive transcriptomic changes in the brain due to T. canis and T. cati infections, demonstrating considerable gene expression shifts related to neuroinflammation and neuronal damage (Waindok et al., 2022). This study shows that murine-derived larvae increase their TcHSP-90 expression in response to findings that parasites in the central nervous system develop stronger stress responses that require heat shock protein activation to survive (Zou et al., 2020). The substantial increase in TcHSP-90 expression at 28 dpi in mice reveals that the parasite faces extreme physical stress when infecting accidental hosts due to the strong immune defense and poor tissue environment provided by these hosts (Yaseen et al., 2020; Abd-Alhassen et al., 2021; Ghazi et al., 2024). Toxocara canis employs complex molecular strategies to alter host immune reactions, which enhances its ability to survive across various host environments (Abd-Alhassen et al., 2021). Macrocyclic lactone resistance in T. canis correlates with changes in drug transporter gene expression, suggesting that the parasite utilizes transcriptional adaptations to overcome environmental stressors (Chelladurai et al., 2020). The upregulation of TcHSP-90 in murine hosts suggests that it may be a part of the general stress-response system of T. canis that enables adaptation to immune-mediated oxidative stress when infecting accidental hosts. The ability of larvae to adapt to various conditions proves essential for their survival in humans because persistent infections involve extended larval movement through tissues and immune system damage. Toxocara canis infection disrupts the IL-33/ST2 pathway, which creates a long-lasting eosinophil-dependent Th2 inflammatory response, leading to an extended infection duration (Raissi et al., 2020). This study confirms that TcCPL-1 expression increases in murine larvae, which probably supports the parasite’s continuous activity and its contact with host immune cells. Therapeutic strategies that target parasite-derived molecules such as ES proteins produced by T. canis are increasingly being investigated. Immune-regulating properties have been demonstrated in these proteins, which present opportunities for new anti-parasitic or immune-modifying therapies (Gao et al., 2022). The results of this study on TcES-1 expression in mice highlight the significance of ES proteins in controlling immune system reactions in hosts. The discovery that other zoonotic nematodes have allergenic genes regulated by gastric conditions indicates potential similar regulatory mechanisms in T. canis that allow parasite-derived factors to dynamically adapt to host environments (Zou et al., 2020; Leal-Silva et al., 2021; Alfatlawi et al., 2021; Klaif et al., 2022; Jesudoss Chelladurai et al., 2023; Etebar et al., 2023). The marked expression variation of TcES-1 and TcCPL-1 genes in mice-derived larvae demonstrates their crucial function in host-parasite communication and presents them as possible therapeutic targets to block parasite immune escape tactics. ConclusionThe results demonstrate that T. canis has developed host-specific transcriptional adaptation mechanisms that allow it to sustain itself in non-permissive hosts while maintaining optimal survival conditions in definitive hosts. Toxocara canis responds to elevated immune pressures in murine hosts by intensifying its molecular defense mechanisms through increased expression of genes that modulate immunity and invade tissues. Understanding the genetic contributors to immune evasion and tissue penetration along with stress response mechanisms establishes the groundwork for subsequent research on targeted therapeutic strategies and diagnostic biomarker development for human toxocariasis. Functional gene validation methods, including RNA interference and CRISPR genome editing, are needed to verify gene functions related to host adaptation and pathogen development. AcknowledgmentThe authors thank the College of Veterinary Medicine at the University of Al-Qadisiyah for their continuous support. Their assistance with laboratory work and guidance throughout this research was greatly appreciated. Conflict of interestThe authors have no conflicts of interest to declare. FundingThe authors have self-funded the study. No external funding source is available. Authors’ contributionsAll authors have participated in the study. Data availabilityData are available when requested by the corresponding author. ReferencesAbd-Alhassen, J.K., Janabi, A.H.D. and Aboktifa, M.A. 2021. Antioxidant and antimicrobial evaluation of lycopene isolated from watermelon. Biochem. Cell. Arch. 21, 2905–2910. Airs, P.M., Brown, C., Gardiner, E., Maciag, L., Adams, J.P. and Morgan, E.R. 2023. WormWatch: park soil surveillance reveals extensive Toxocara contamination across the UK and Ireland. Vet. Rec. 192(1), 1. Alfatlawi, M.A., Jasim, A.A., Jarad, N.E. and Khlaif, S.F. 2021. Clinical and molecular identification of circulating Theileria annulata strains in calves in Al-Diwaniyah, Iraq. Iraqi. J. Vet. Sci. 35(1), 115–119. Cabral Monica, T., Evers, F., Souza Lima Nino, B., Pinto‐Ferreira, F., Breganó, J.W., Ragassi Urbano, M., Rubinsky‐Elefant, G., Freire, R.L., Navarro, I.T. and Mitsuka‐Breganó, R. 2022. Socioeconomic factors associated with infection by Toxoplasma gondii and Toxocara canis in children. Transbound. Emerg. Dis. 69(3), 1589–1595. Dar, Z.A., Syed, T. and Yattoo, G.N. 2016. Toxocariasis in Kashmir: epidemiology, pathogenesis and immunological studies in human population. Srinagar, India: University of Kashmir. Etebar, F., Hosseini, S.H., Borhani Zarandi, M., Moghadasi, A.N. and Jalousian, F. 2023. Immunomodulatory effects of C-type lectin protein of Toxocara canis on experimental autoimmune encephalomyelitis. Parasite Immunol. 45(11), e13010. Gao, J., Zou, Y., Wu, X.J., Xu, Y., Zhu, X.Q. and Zheng, W.B. 2022. Differential miRNA expression profiles in the bone marrow of Beagle dogs at different stages of Toxocara canis infection. BMC Genomics 23(1), 847; doi: 10.1186/s12864-022-09081-8 Gelmi-Candusso, T.A., Wheeldon, T.J., Patterson, B.R. and Fortin, M.J. 2024. The effect of urbanization and behavioral factors on coyote net displacement and its implications for seed dispersal. Urban Ecosyst. 27(2), 387–397. Ghazi, A.M., Ali Al-bayati, M.A. and Janabi, A.H.D. 2024. Metabolomics-detected alterations generated by phytosomal propolis and phytosomal lycopene in male rats with induced BPH. Iraqi. J. Vet. Sci. 38(Suppl. I-IV), 7–15. Henke, K., Ntovas, S., Xourgia, E., Exadaktylos, A.K., Klukowska-Rötzler, J. and Ziaka, M. 2023. A semi-systematic review on the enduring impact of toxocariasis. Int. J. Environ. Res. Public. Health. 20(21), 6972. Holland, C.V. 2023. A walk on the wild side: a review of the epidemiology of Toxocara canis and Toxocara cati in wild hosts. Int. J. Parasitol. Parasites Wildl. 22, 216–228. Jahanmahin, A. and Borji, H. 2023. Unveiling the zoonotic significance of toxocariasis in humans: the role of Toxocara canis. Small. Anim. Adv. 2, 5–11 Jesudoss Chelladurai, J.R.J., Jones, D.E. and Brewer, M.T. 2021. Characterization of a P-glycoprotein drug transporter from Toxocara canis with a novel pharmacological profile. Int. J. Parasitol. Drugs Drug Resist. 17, 191–203; doi: 10.1016/j.ijpddr.2021.10.002 Jesudoss Chelladurai, J.R.J., Martin, K.A., Vardaxis, P., Reinemeyer, C., Vijayapalani, P., Robertson, A.P. and Brewer, M.T. 2023. Repertoire of P-glycoprotein drug transporters in the zoonotic nematode Toxocara canis. Sci. Rep. 13, 4971; doi: 10.1038/s41598-023-31556-1 Klaif, S.F., Jassim, A., Alfatlawi, M.A. and Ali, M.J. 2022. Major-surface-protein-4-gene-based detection of Anaplasma marginale in sheep in Al-Diwaniyah, Iraq. Iraqi J. Vet. Sci. 36(1), 85–88. Leal-Silva, T., Vieira-Santos, F., Oliveira, F.M.S., Padrão, L.L.S., Kraemer, L., Matias, P.H. da P., Lopes, C. de A., Ruas, A.C.L., Azevedo, I.C. de, Nogueira, D.S., Rachid, M.A., Caliari, M.V., Russo, R.C., Fujiwara, R.T. and Bueno, L.L. 2021. Detrimental role of IL-33/ST2 pathway sustaining a chronic eosinophil-dependent Th2 inflammatory response, tissue damage and parasite burden during Toxocara canis infection in mice. PLoS Neglected Trop. Dis. 15(7), e0009639; doi:10.1371/journal.pntd.0009639 Magnaval, J.F., Bouhsira, E. and Fillaux, J. 2022. Therapy and prevention for human toxocariasis. Microorganisms. 10, 241–260. Qualizza, R., Incorvaia, C., Grande, R. and Makri, E. et al. 2011. Seroprevalence of IgG anti-Toxocara species antibodies in patients with suspected allergy. Auckland, New Zealand: Dove Medical Press. Raissi, V., Mohebali, M., Kia, E.B., Rahimi Foroushani, A., Sohrabi, N., Rokni, M.B. and Zibaei, M. 2020. Expression of miR-21 and miR-103a in Toxocara canis: potential diagnostic markers. Iranian J. Parasitol. 15(4), 559–567. Saelens, G., Robertson, L. and Gabriël, S. 2022. Diagnostic tools for the detection of taeniid eggs in different environmental matrices: a systematic review. Food Waterborne Parasitol. 26, e00143–e00152. Saichenko, I.V., Antipov, A.A., Bakhur, T.I., Shmayun, S.S. and Bezditko, L.V. 2021. Co-infection of Trichuris vulpis and Toxocara canis in different aged dogs: influence on haematological indices. Biosyst. Divers. 29(2), 129–134. Schwartz, R., Bidaisee, S., Fields, P.J., Macpherson, M.L. et al. 2021. The epidemiology and control of Toxocara canis in puppies. Amsterdam, The Netherlands: Elsevier Ltd. Schwartz, R., Bidaisee, S., Fields, P.J., Macpherson, M.L.A. and Macpherson, C.N.L. 2022. The epidemiology and control of Toxocara canis in puppies. Parasite Epidemiol. Control 16, 232. Tylkowska, A., Mocha, N., Kołnierzak, M.M. and Szenejko, M. 2024. Risk factors associated with soil-transmitted helminths in dog feces contaminating public areas of Warsaw, Poland. Animals. 14, 450–461. Waindok, P., Janecek-Erfurth, E., Lindenwald, D.L., Wilk, E., Schughart, K., Geffers, R. and Strube, C. 2022. Toxocara canis- and Toxocara cati-induced neurotoxocarosis is associated with comprehensive brain transcriptomic alterations. Microorganisms 10(1), 177; doi: 10.3390/microorganisms10010177 Xu, Y., Zheng, W.B., Li, H.Y., Cai, L., Zou, Y., Xie, S.C., Zhu, X.Q. and Elsheikha, H.M. 2022. RNA sequencing reveals dynamic expression of spleen lncRNAs and mRNAs in Beagle dogs infected by Toxocara canis. Parasites Vectors 15(1), 279; doi: 10.1186/s13071-022-05380-x Yaseen, M.M., Karawan, A.C., Alfatlawi, M.A.A. and Janabi, A.H.D. 2020. Gut bacterial cytochrome-P450 of mosquito larvae in degradation of temephos insecticide. Ann. Trop. Med. Public Health 23(1), S412. Zhu, X.Q., Korhonen, P.K., Cai, H. and Young, N.D. et al. 2015. Genetic blueprint of the zoonotic pathogen Toxocara canis. London, United Kingdom: Nature Publishing Group / Springer Nature. Zou, Y., Zheng, W.B., Elsheikha, H.M., He, J.J., Lu, Y.X., Wang, S., Guo, A. and Zhu, X.Q. 2023. Modulation of long noncoding RNA (lncRNA) and messenger RNA (mRNA) expression in the liver of Beagle dogs by Toxocara canis infection. Parasites Vectors 16(1), 114; doi: 10.1186/s13071-023-05738-9 Zou, Y., Zheng, W.B., He, J.J., Elsheikha, H.M., Zhu, X.Q. and Lu, Y.X. 2020. Toxocara canis affects hepatic microRNA expression in Beagle dogs at different infection stages. Front. Vet. Sci. 7, 587273. | ||
| How to Cite this Article |
| Pubmed Style Al_shabbani AHA, Al-kaeebi SRA, Madhloom MH, Alfatlawi MAA. Stage-specific gene expression profiling of Toxocara canis in canine and human hosts: Implications for the pathogenesis and transmission of zoonotic diseases. Open Vet. J.. 2026; 16(5): 2970-2978. doi:10.5455/OVJ.2026.v16.i5.40 Web Style Al_shabbani AHA, Al-kaeebi SRA, Madhloom MH, Alfatlawi MAA. Stage-specific gene expression profiling of Toxocara canis in canine and human hosts: Implications for the pathogenesis and transmission of zoonotic diseases. https://www.openveterinaryjournal.com/?mno=297338 [Access: June 26, 2026]. doi:10.5455/OVJ.2026.v16.i5.40 AMA (American Medical Association) Style Al_shabbani AHA, Al-kaeebi SRA, Madhloom MH, Alfatlawi MAA. Stage-specific gene expression profiling of Toxocara canis in canine and human hosts: Implications for the pathogenesis and transmission of zoonotic diseases. Open Vet. J.. 2026; 16(5): 2970-2978. doi:10.5455/OVJ.2026.v16.i5.40 Vancouver/ICMJE Style Al_shabbani AHA, Al-kaeebi SRA, Madhloom MH, Alfatlawi MAA. Stage-specific gene expression profiling of Toxocara canis in canine and human hosts: Implications for the pathogenesis and transmission of zoonotic diseases. Open Vet. J.. (2026), [cited June 26, 2026]; 16(5): 2970-2978. doi:10.5455/OVJ.2026.v16.i5.40 Harvard Style Al_shabbani, A. H. A., Al-kaeebi, . S. R. A., Madhloom, . M. H. & Alfatlawi, . M. A. A. (2026) Stage-specific gene expression profiling of Toxocara canis in canine and human hosts: Implications for the pathogenesis and transmission of zoonotic diseases. Open Vet. J., 16 (5), 2970-2978. doi:10.5455/OVJ.2026.v16.i5.40 Turabian Style Al_shabbani, Amal Hassan Abd, Saafa Ressan Abdullah Al-kaeebi, Miras Hasan Madhloom, and Monyer Abdulamier Abd Alfatlawi. 2026. Stage-specific gene expression profiling of Toxocara canis in canine and human hosts: Implications for the pathogenesis and transmission of zoonotic diseases. Open Veterinary Journal, 16 (5), 2970-2978. doi:10.5455/OVJ.2026.v16.i5.40 Chicago Style Al_shabbani, Amal Hassan Abd, Saafa Ressan Abdullah Al-kaeebi, Miras Hasan Madhloom, and Monyer Abdulamier Abd Alfatlawi. "Stage-specific gene expression profiling of Toxocara canis in canine and human hosts: Implications for the pathogenesis and transmission of zoonotic diseases." Open Veterinary Journal 16 (2026), 2970-2978. doi:10.5455/OVJ.2026.v16.i5.40 MLA (The Modern Language Association) Style Al_shabbani, Amal Hassan Abd, Saafa Ressan Abdullah Al-kaeebi, Miras Hasan Madhloom, and Monyer Abdulamier Abd Alfatlawi. "Stage-specific gene expression profiling of Toxocara canis in canine and human hosts: Implications for the pathogenesis and transmission of zoonotic diseases." Open Veterinary Journal 16.5 (2026), 2970-2978. Print. doi:10.5455/OVJ.2026.v16.i5.40 APA (American Psychological Association) Style Al_shabbani, A. H. A., Al-kaeebi, . S. R. A., Madhloom, . M. H. & Alfatlawi, . M. A. A. (2026) Stage-specific gene expression profiling of Toxocara canis in canine and human hosts: Implications for the pathogenesis and transmission of zoonotic diseases. Open Veterinary Journal, 16 (5), 2970-2978. doi:10.5455/OVJ.2026.v16.i5.40 |

