




| Research Article | ||
Open Vet. J.. 2026; 16(5): 3203-3217 Open Veterinary Journal, (2026), Vol. 16(5): 3203-3217 Research Article Mucosal delivery of pleurotus-based ISCOMATRIX elicits humoral and cellular immunity against pneumococcal infectionAkunna Perpetua Emeruwa1,2,3*, Angus Nnamdi Oli2, Malachy Chigozie Ugwu2,4, Adekunle B. Rowaiye5and Chinemerem Paschaline Ogbu61Department of Microbiology and Parasitology, Faculty of Allied Health Sciences, David Umahi Federal University of Health Sciences, Uburu, Nigeria 2Department of Pharmaceutical Microbiology and Biotechnology, Faculty of Pharmaceutical Sciences, Agulu, Nnamdi Azikiwe University, Awka, Nigeria 3International Institute for Pharmaceutical Research and Innovation, David Umahi Federal University of Health Sciences, Uburu, Nigeria 4Nnamdi Azikiwe University Tetfund Center of Excellence for Biomedical, Engineering and Agricultural Translational Studies, Awka, Nigeria 5Departament of Pharmaceutical Sciences, North Carolina Central University, Durham, USA 6Department of Biochemistry, University of Nebraska-Lincoln, Lincoln, NE 68588, USA *Corresponding Author: Akunna Perpetua Emeruwa. Department of Microbiology and Parasitology, Faculty of Allied Health Sciences, David Umahi Federal University of Health Sciences, Uburu, Nigeria. Email: perpetuanwigwe [at] gmail.com Submitted: 08/10/2025 Revised: 30/03/2026 Accepted: 11/04/2026 Published: 31/05/2026 © 2025 Open Veterinary Journal
AbstractBackground: Pleurotus ostreatus is an edible mushroom with known immunomodulatory compounds, such as saponins, with potential for vaccine adjuvant development. Aim: This study investigated the immunogenicity of an ISCOMATRIX-based adjuvant made with Pleurotus-derived saponins and delivered via mucosal routes to elicit protective immune responses against Streptococcus pneumoniae infection. Methods: Saponins were extracted from P. ostreatus using aqueous extraction, defatted with ethyl acetate, and purified using vacuum liquid chromatography. The ISCOMATRIX formulation was prepared by lipid film hydration and characterized accordingly. Mice were immunized through intranasal, oral, or rectal routes and evaluated for delayed-type hypersensitivity, bacterial load, neutrophil recruitment, antibody titers [Immunoglobulin A (IgA), Immunoglobulin G IgG)], and cytokine levels (Interferon ᵞ, Interleukin 4) using standard assays and enzyme-linked immunosorbent assay. Results: Mice immunized intranasally and rectally showed significantly reduced inflammation and bacterial counts compared with controls (p < 0.05). Neutrophil recruitment was notably increased in the nasal and oral groups (p < 0.05), whereas rectal administration was not statistically significant (p > 0.05). All mucosal routes induced significantly elevated IgA and IgG titers, particularly via the intranasal route (p < 0.001). Elevated measures of Interferon-γ and Interleukin-4 were observed in splenocyte cultures from all immunized groups, indicating both Th1 and Th2 activation. Conclusion: Mucosal delivery of Pleurotus-derived saponin ISCOMATRIX formulations can induce robust immune responses against S. pneumoniae in animal models. Intranasal administration showed the most consistent immunogenicity among the tested routes. This platform holds potential as a non-invasive alternative to injectable pneumococcal vaccines, warranting further preclinical development. Keywords: Adjuvant, Iscomatrix, Mucous organs, Mushroom, Streptococcus. IntroductionStreptococcus pneumoniae remains a leading contributor to global illness and death, with its heaviest toll seen among children aged 5 years and older adults (Centers for Disease Control and Prevention , 2019). Data from the World Health Organization and the Nigeria Center for Disease Control indicate that pneumococcal disease accounts for over 1 million deaths annually, particularly in low- and middle-income countries where it continues to drive high childhood mortality (Centers for Disease Control and Prevention, 2014; World Health Organization, 2019). Despite the introduction of effective pneumococcal vaccines, this pathogen persists as the predominant cause of community-acquired pneumonia, manifesting in severe invasive diseases such as meningitis and bacteremia, as well as in non-invasive conditions like otitis media, sinus infections, and localized pneumonia (Musher and Thorner, 2014; Malihe et al., 2020). Pneumococcus asymptomatically colonizes the nasopharynx, making carriers an important reservoir for community-wide transmission (Chao et al., 2014). Conventional vaccines, while effective, face notable limitations, including the need for skilled personnel for administration, multiple dosing schedules, reduced mucosal immunogenicity, and needle-phobia and compliance issues. A pilot study in Anambra State, Nigeria, revealed that 84% of unvaccinated individuals resisted vaccination due to a fear of injections and lack of trained personnel (Unpublished doctoral dissertation). In some cases, infection occurred despite vaccination, which was attributed to incomplete dosing and suboptimal immune responses. Traditional injectable vaccines are often inadequate at inducing robust mucosal immunity, which is crucial because many pathogens, including S. pneumoniae, enter the host through mucosal surfaces (Su et al., 2016). In addition, injectable routes carry risks of needle-stick injuries and bloodborne disease transmission. Antigen delivery, which includes both the delivery of the antigen to provide the required signals for the activation and maturation of pertinent antigen-presenting cells (APCs) and the administration of drugs to specific sites of the body, is a critical component in addressing the challenges involved in vaccine development (Obaji et al., 2025). Report has shown that vaccine administration through the mucosal organs can produce systemic immunity in a manner similar to that of needle and syringe immunization (Obaji et al., 2025). Most viral and bacterial infections start at the mucosal surfaces, where they invade the host or establish infection. Antigen delivery to these surfaces can mimic natural infection, prompting local and systemic immune responses. Given these challenges, mucosal vaccine delivery systems that mimic natural infection routes and stimulate immune responses both locally and systemically have attracted increasing interest (Brito et al., 2013). Saponins—especially those derived from Quillaja saponaria, such as Quil A—have long been recognized as potent vaccine adjuvants, largely because of their ability to induce Th1-mediated and cytotoxic T lymphocyte immune responses (Kensil et al., 1991; Sun et al., 2009). In recent years, they have been developed into Immunostimulatory Complexes (ISCOMs), which are particulate carriers composed of saponins, cholesterol, and phospholipids (Lövgren et al., 2011). These nanostructures enhance antigen uptake, promote efficient presentation, and elicit robust humoral and cellular immune responses while minimizing toxicity (Sun et al., 2009; Lövgren et al., 2011). ISCOMATRIX, an antigen-free derivative of ISCOMs, preserves the same architecture and immunostimulatory properties as ISCOMs (Rowaiye et al., 2022). The overall efficacy of these systems depends on factors such as particle size, surface charge, and their interaction with APCs (Behboudi et al., 1996; Obaji et al., 2025). ISCOMs can stimulate all classes of immunoglobulins, robust T-cell responses, and delayed-type hypersensitivity, making them promising tools for both parenteral and mucosal immunization. In recent years, interest in exploring natural adjuvants beyond Q. saponaria has grown. Edible mushrooms, particularly those in the Pleurotus genus (P. ostreatus, or oyster mushrooms), have been widely studied for their nutritional, immunomodulatory, and antimicrobial effects (Chang and Miles, 1992; Wang and Ng, 2000; Ngai and Ng, 2004). These macrofungi are rich in proteins, vitamins, fiber, and secondary metabolites, such as sterols, terpenoids, flavonoids, and, notably, saponins (Garcia-Lafuente et al., 2011). Pleurotus ostreatus also contains antioxidants and enzymes capable of breaking down lignin and phenolic compounds (Fountoulakis et al., 2002), and it can suppress pro-inflammatory cytokines, e.g., tumor necrosis factor-alpha (Rowaiye et al., 2022). It also exhibits phagocytic activity against S. pneumoniae infections (Oli et al., 2019). Pleurotus ostreatus is a promising source for novel adjuvant development given its saponin content and immune-enhancing potential. However, while ISCOMATRIX systems have been well studied using Q. saponaria saponins, data on the immunogenic potential of ISCOMATRIX derived from P. ostreatus—particularly when administered via mucosal routes—are limited. According to Oli et al. (2019), the immunostimulating properties of P. ostreatus have been studied, but there is no data on the use of P. ostreatus or its ISCOMATRIX directives as adjuvants in vaccine development. This forms the basis for this study. This study aimed to evaluate the immune-enhancing capability of an ISCOMATRIX adjuvant formulated with Pleurotus-derived saponins and delivered through intranasal, oral, and rectal routes to protect against S. pneumoniae. This study uses a vaccine delivery system that targets mucosal surfaces, offering an alternative to injectable pneumococcal vaccines. By enhancing antigen stability and immunogenicity, mucosal administration aims to leverage the body's largest immunological organ: the mucous membranes. This study specifically investigates the ability of mucosal vaccines to elicit both cellular and humoral responses to advance the development of needle-free, mucosal vaccines for pneumococcal diseases. Materials and MethodsMushroom selection and identificationFresh fruiting bodies of P. ostreatus (3 kg) were collected from Dilomat Farm in Port Harcourt, Rivers State, Nigeria. The species was taxonomically described and authenticated by a mycologist at the Department of Pharmacognosy and Traditional Medicine, Faculty of Pharmaceutical Sciences, Nnamdi Azikiwe University, Agulu, Nigeria. A voucher specimen (Voucher No: PLG/474/A/038) was deposited for future reference in the departmental herbarium. Pleurotus ostreatus aqueous extract preparationThe mushrooms were thoroughly washed with distilled water to remove dirt, air-dried at ambient conditions of 25°C ± 2°C for 7 days, and subsequently milled into a fine powder using a mechanical grinder fitted with a 1 mm mesh sieve. Then, 2 kg of powdered mushroom was weighed, transferred to a beaker containing 800 ml of distilled water, and mixed with a magnetic stirrer for 8 hours. The sample suspension in a beaker was placed on a hot plate maintained at 70°C ± 4°C and heated for 4 hours with intermittent stirring. The mixture was then cooled to 45°C and filtered three times through a Buchner funnel fitted with two layers of Whatman No. 4 filter paper using a 1 l suction flask under vacuum to remove retained residues, thereby improving the extract yield. The filtered extract was treated with 50 ml of 2% gelatin solution to precipitate condensed tannins. After precipitation, the extract was subjected to vacuum filtration using a Buchner funnel lined with three sheets of qualitative filter paper and connected to a 1 l suction flask. To obtain crude saponin, the defatting method described by Christopher (2017) was performed. Defatting was performed to remove fats and lipids that may interfere with the extraction/purification process. Briefly, the extract was transferred into a 2-l separatory funnel and partitioned with 200 ml of ethyl acetate, then left for a few minutes to settle. The ethyl acetate phase was discarded. Finally, the extract was concentrated in a rotary evaporator using a round-bottom flask at a temperature below 45°C, and the resulting residue was stored at 4°C under refrigeration. Qualitative determination of saponinA frothing test was performed as described by Jeffrey and Harborne (1973) and modified by Oli et al. (2019) to further identify the saponins. The extract (0.5 g) was diluted with 20 ml of distilled water and continuously vortexed in a graduated cylinder for 15 minutes. The foam height (which lasted for 5 minutes) was measured. Góral and Wojciechowski (2020) described saponins as secondary amphiphilic glycosidic metabolites with foaming properties produced by plants. Therefore, the production of a 1-cm foam layer was indicative of the presence of saponins (Góral and Wojciechowski, 2020). The frothing test was conducted in triplicate to eliminate inefficiency. Purification of saponin by column chromatographyMaterials and preparation of columnsThe saponin purification column chromatography method described by Negi et al. (2011) was performed. A 100 g sample of C-18 reverse-phase silica gel was weighed and suspended in 96% ethanol (1.5 l). The resulting suspension was poured into a measuring glass column and allowed to settle into a stationary phase. The column was gently tapped intermittently to remove air bubbles, allowing the silica to pack tightly into the column. After packing, rinsing of the column was performed by slowly pipetting solvent down the inner wall to remove any loosely packed particles. The solvent was then carefully drained until its level was flush with the stationary phase surface. Sample loadingThen, 100 ml of distilled water was added and drained again to equilibrate the stationary phase. A 1 g aqueous extract residue containing saponin was dissolved in 10 ml of distilled water and loaded onto a silica gel column using a Pasteur pipette. A small cotton plug was placed over the column loading site to prevent sample leakage. Elution and fraction collectionElution was performed in a stepwise methanol-water gradient starting from 100% distilled water to 100% methanol, with the methanol concentration increasing by 10% per step. At each step, 100 ml of solvent was passed through the column while the eluate fractions were collected under vacuum using a suction flask. A total yield of 9 fractions of 100 ml each was obtained. Thin-layer chromatography was further used to analyze the fractions for saponin content. The fractions with the highest saponin content were further eluted with 90% methanol and dried in a rotary evaporator using a round-bottom flask at a temperature below 45°C. Concentrated pure saponin extract was stored in an airtight container at room temperature away from humidity. Column cleaningThe column was washed with 100 ml of 100% ethanol, followed by 100 ml of 70% ethanol, and finally 100 ml of distilled water. Each solvent was allowed to stand in the stationary phase for 10 minutes before being drained to restore the column. Formulation of the iscomatrixThe lipid film hydration method, as described by Jafar et al. (2018), was employed in the iscomatrix formulation. Briefly, 2:2:1 (w/w/w) ratios of pure egg phosphatidylcholine lipid: Pleurotus saponin: cholesterol (≥93%) were used. First, 8 mg of EPC and 4 mg of cholesterol were dissolved in chloroform (≥96.06% purity). The solvent (chloroform) was removed under reduced pressure using a rotary evaporator (Hettich, Germany). This was done to produce a thin, dried, and uniform lipid film. Subsequently, a 1:1 (v/v) mixture of tert-butanol and sucrose solution (100 mg/ml) was added to the dried lipid film (4 ml total) and stirred for 5 minutes. The obtained lipid phase was flash-frozen on dry ice and lyophilized overnight at 60°C. A phosphate buffered saline (PBS) solution (0.01 M, pH 7.4) of Plerotus saponin (8 mg/4 ml) was poured into the lipid phase to hydrate the solid matrices. The hydrated mixture was sonicated in a water bath at 40 kHz for 10 minutes to ensure uniformity and homogeneity of iscomatrix nanoparticles to promote dispersion. The resulting suspension was then extruded through polycarbonate membranes with pore sizes ranging from 100 to 400 nm, and the resulting iscomatrix samples were stored at 4°C in airtight amber glass vials. Characterization of the iscomatrixThe morphology and structure of the iscomatrix were determined using a transmission electron microscope (Phillips CM100, Philips Electron Optics, Holland) following the method of Titus et al. (1985). In a nutshell, samples were placed on carbon-coated, glow-discharged copper grids and negatively stained with 2% phosphotungstic acid (pH 5.2). Subsequently, an electron microscope operating at a 100 kV acceleration voltage and 93,000× magnification was used to scan the samples. Using a Zetasizer (Nano-ZS, Malvern Instruments, UK), the zeta potential and size of formulations and the Polydispersity index (PDI) were ascertained by following the procedure outlined by Jaafari et al. (2007). The iscomatrix samples were further characterized by dispersing them in distilled water at a concentration of 1 mg/ml to obtain the following: • Solubility: One milligram of iscomatrix was diluted in 1 milliliter of distilled water to ascertain the solubility level. • Determination of Ph: A pH meter (Genlab, UK) was used to determine the pH of the iscomatrix at room temperature. Inactivation of S. pneumoniaeThe previously stored human invasive and non-invasive pneumococcal serotypes (6A, 19F, 23F, 19A, and 18C) were used for immunization and challenge in the Department of Pharmaceutical Microbiology, Nnamdi Azikiwe University Awka, Anambra State. A 0.5 McFarland standard of each pneumococcus serotype (1.5 × 10⁸ CFU/ml) was cultured in Todd–Hewitt broth (Ken Light Laboratory, Mumbai, India) containing 17% fetal calf serum (Sigma-Aldrich, USA). Cultures were incubated for 18 hours at 36°C in 5% CO₂. Bacterial cells were collected by centrifugation and washed three times with sterile pyrogen-free saline to obtain heat-killed organisms for immunization (Aaberge and Løvik, 1996). To determine the bacterial load, 10-fold serial dilutions (0.1 ml) were spread onto horse blood agar plates. After incubation at 37°C for 36 hours, viable counts were calculated based on colony numbers, using an untreated suspension as the control. Animal modelThe animals used in this research were obtained from the Department of Zoology, Nnamdi Azikiwe University, Awka. Animal studies were approved by the Institutional Animal Care and Use Committee at Nnamdi Azikiwe University (IACUC approval number: NAU/AREC/2023/000110). A total of 60 male and female BALB/c mice aged between 2 and 3 months and weighing 20–30g, were included. Eight to ten animals per cage were kept in a room with a 12:12 light-dark cycle, controlled humidity (40%–70%), and temperature (23°C ± 1°C). Water and food were freely available. All mice were cared for under standard guidelines, and a 1-week acclimatization period in a pathogen-free environment was provided before the start of experiments. Preclinical screening of the safety of Iscomatrix pneumococcal vaccinesToxicity testingThis was calculated using the approach described by Garrido-Acosta et al. (2014). This approach involved two stages, involving seventeen animals in both. Nine animals, each divided into three groups, were used in the first phase. Different doses of the test vaccine candidate (10, 100, and 1000 mg/kg) were administered to the animals. After that, the animals were observed for a whole day to track their behavior and possible death. For the second stage, eight animals—four groups of two animals each—were used. Higher doses of the test drug (2,000, 3,000, 4,000, and 5,000 mg/kg) were administered to the animals, and they were observed for 24 hours for behavioral changes and mortality. The LD50 was determined using the following formula: LD50=√ (D0 X D100) where D0 represents the highest dose that resulted in no mortality, and D100 represents the lowest dose that produced mortality. Animal body weight changes testThis was performed using the method described by Haruka et al. (2010) with modifications from Oli et al. (2015). A total of 50 animals with related weight values were used. Animals were divided into five groups of 10 animals per group. Group 1 had weight values of 20–25g; group 2 weight ranged from 24.5–25.5g; group 3 had a weight range was 22–23.5g; group 4 had a weight range of 23.5–25g and group 5 weight ranged from 23–24g. The animals were weighed on day 0 before the vaccine candidate was administered. Each group of animals was anesthetized using methoxyflurane by inhalation, then saline (20 mg) was administered to the control group, and the iscomatrix vaccine candidate was administered orally at doses of 10, 50, 100, and 500 mg/group. The animals were then placed under observation while the weight gain/loss was measured on days 3 and 7. The difference in weight was compared with that of the control group on day 0. According to Kuarta (2006), a good vaccine should not: (i) result in the mice losing weight 3 days after the injection; (ii) cause the mice to gain 60% of their initial weight compared to the control group 7 days after the injection; (iii) cause more than 5% of the animals to die during the 7-day observation period; or (iv) show any signs of illness in the animals (Kuarta, 2006). Before and after administration, the body weight (BW) was measured, and the difference was computed. ImmunizationFive mouse groups (one to five), each with six mice, received two immunizations spaced 3 weeks apart at three distinct mucosal sites (oral, rectal, and intranasal). 50 mg of iscomatrix was suspended in 0.2 ml of PBS for immunization. Mice were anesthetized using methoxyflurane administered by inhalation. Mice in different groups were immunized with the following formulations: Group 1: 50 mg iscomatrix vaccine candidate + 0.05 mg inactivated pneumococcus / animal intranasally, Group 2: 50 mg iscomatrix vaccine candidate + 0.05 mg inactivated pneumococcus/animal orally, Group 3: 50 mg iscomatrix vaccine candidate + 0.05 mg inactivated pneumococcus/animal rectally, Group 4: (500 mg pneumococcal conjugate vaccine + 0.05 mg inactivated pneumococcus/animal intramuscularly as positive control, and Group 5: (saline as negative control. The mouse was placed in a supine position with its head down for intranasal immunization. The vaccine was gently injected into the nares using a micropipette to allow the mouse to sniff it in. Using a micropipette, the vaccine was gradually administered for oral immunizations, allowing the mouse to suck the fluid from the tip. A feeding tube was used to administer the rectal immunization vaccine. The tube was introduced into the anus, with the tip placed approximately 3 cm from the anal entrance. Challenge with PneumococcusTwo weeks after the final booster administration, a 0.5 McFarland standard of pneumococcus (1.5 × 10^8) overnight broth in a 50 μl volume was injected subcutaneously into the left footpad of both the immunized and control groups. The thickness of the left footpad of the animals was measured using a digital caliper at weekly intervals for 6 weeks and compared with the thickness before infection (that is, the delayed hypersensitivity reaction). Quantitative determination of the burden after the challengeThe quantitation of bacterial load in the blood of infected animals was performed by plating 25-µl aliquots of serial 10-fold dilutions onto agar plates. Plates were incubated at 36°C under 5% CO₂ for 18 hours, after which CFUs were counted. The spleen and footpad were harvested from each experimental group. Viable pneumococcal organisms in the spleen and footpad of mice were quantified using the limiting dilution assay described by Jaafari et al. (2007). Briefly, mice were sacrificed at week 8 post-challenge, and the feet were aseptically excised and homogenized in RPMI-1640 (Titan Biotech Ltd.) supplemented with 10% (v/v) heat-inactivated fetal calf serum (FCS), 2 mM glutamine, 100 U/ml penicillin, and 100 µg/ml streptomycin sulfate (RPMI-FCS). The homogenates were serially diluted (8-fold, 10-fold dilutions) and inoculated into flat-bottom 96-well microtiter plates containing a solid rabbit blood agar layer (tetraplicate wells). Plates were incubated at 25°C ± 1°C for 7–10 days, after which the presence or absence of motile bacteria was examined using an inverted microscope. Viable counts were estimated using the GraphPad Prism software, a statistical model for a limiting dilution assay. Blood collectionBlood samples (250 μl) were obtained from the orbital venous plexus of mice under light CO₂ anesthesia and collected in EDTA-coated microcentrifuge tubes. The samples were gently mixed and processed within 1 hour of collection using an automated hematology analyzer. Manual determination of the neutrophil countNeutrophil counts were manually determined from blood smears prepared using EDTA-treated samples. Smears were air-dried and stained with Hemacolor Stain for Microscopy. The slides were examined using light microscopy (Olympus, UK), and 150–200 leukocytes were counted. The EasyCell counting software was used to eliminate the inefficiencies associated with mechanical desktop tally counters. Leukocytes were categorized into the following groups: eosinophils, basophils, segmented neutrophils, band neutrophils, lymphocytes, monocytes, and other cells. The percentage for each category was calculated by dividing the total number of leukocytes counted by the number of cells in each group. By dividing the total number of leukocytes counted by the number of cells in each group, the percentage for each category was calculated. Triplicate counts were performed on 12 of the sixty slides to determine the counting procedure’s accuracy. Calculation of the absolute neutrophil count (ANC)The total leukocyte count white blood cell (WBC) multiplied by the percentages of segmented neutrophils (Ns) and band neutrophils (Nb) yielded the manual ANC (ANCm) as follows: ANCm=WBC (cells/µl) × (percent (Ns + Nb) +100). Quantitation of the total antibody assayMice were bled retro-orbitally before and 10 days after the challenge to obtain blood samples. After allowing the blood to coagulate at 4°C, the serum was extracted using a 10-minute centrifugation at 14,000 rpm. Serum samples were stored at -20°C. Total Immunoglobulin G (IgG) and Immunoglobulin A (IgA) antibody titration and assessment were performed using an ELISA technique described by Badiee et al. (2007). Briefly, 96-well plates were coated with 50 µl of pneumococcal antigen (10 μg/ml) in phosphate-buffered saline (pH 7.3) and kept overnight at 4°C. The plates were blocked with 300 μl of 2.5% bovine serum albumin in PBS-Tween in each well for 1 hour at 37°C after being cleaned with PBS-Tween solution. This is done to stop the antigen and antibody from binding to each other non-specifically. The serum was added to the plates at various dilutions (1:200, 1:2,000, 1:20,000, and 1:200,000) and allowed to sit for 75 minutes at 37°C for 75 minutes. Horseradish peroxidase-labeled rabbit anti-mouse IgG and IgA secondary antibodies were used to detect the antibodies following the manufacturer’s instructions. Quantitation of cytokine levels in culture supernatantsThe concentrations of Interleukin 4 (IL-4) and Interferon ᵞ (IFN-γ) in culture supernatants were quantified using ELISA, following the protocol of Badiee et al. (2007) with some modifications. At week 3 after the final booster, three mice per group were sacrificed concurrently with the challenge experiment. The spleens were aseptically excised, and mononuclear cells were isolated by Ficoll–Hypaque density gradient centrifugation, as described by Taswell (1981). The cells were then washed and resuspended in RPMI 1640-FCS medium, and seeded at a concentration of 2 × 10^6 cells/ml in 96-well flat-bottom plates. Next, the spleen cells were re-stimulated for 3 days with pneumococcal antigen (10 µg/ml) and incubated at 37°C with 5% CO2 for 72 hours. Subsequently, the culture supernatants were harvested, and the levels of IL-4 and IFN-γ cytokines were measured by capture enzyme-linked immunosorbent assay using commercial kits, following the manufacturers’ instructions. Statistical analysisStatistical analyses were performed using IBM SPSS Statistics version 25 (IBM Corp., USA) alongside Microsoft Excel 2010. Differences among groups were analyzed using one-way analysis of variance. Where analysis of variance revealed significant variation (p ≤ 0.05), Tukey–Kramer post hoc testing was applied to determine specific group differences. The results are reported as mean values accompanied by standard deviations. A p-value threshold of ≤ 0.05 was set to denote statistical significance, with levels represented as *p < 0.05, **p < 0.01, and ***p < 0.001. Ethical approvalInstitutional Animal Care and Use Committee at Nnamdi Azikiwe University/IACUC approval number: NAU/AREC/2023/000110. Ethical approval date is 23rd October, 2023. ResultsCrude saponin isolationFigure 1 shows the image of the fresh P. ostreatus from which saponin was isolated. Powdered P. ostreatus (2 kg) yielded 20% crude saponin after isolation. Confirmation of saponin was by the formation of a foam layer during a frothing test. Purification of the crude saponin yielded the pure sample used for iscomatrix formulation.
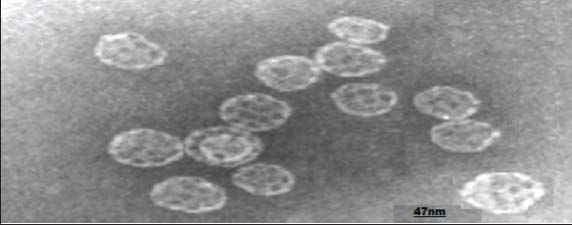
Fig. 1. Pleurotus ostreatus. Characteristics of the formulated IscomatrixThe formulated iscomatrix has a size, zeta potential, and PDI equivalent to those of the standard iscomatrix already studied. These characteristics are presented in Table 1. Transmission electron microscopy (TEM) revealed the morphology of a hollow cage-like colloidal structure (Fig. 2). Table 1. Characterization of Iscomatrix.
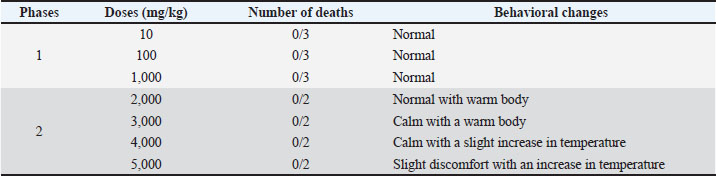
Fig. 2. Iscomatrix particle size and morphology. Acute toxicityThe iscomatrix vaccine candidate, administered at doses of 10, 100, 1,000, 2,000, 3,000, 4,000, and 5,000 mg/kg BW in both the first and second phases of the toxicity study, showed no lethality. However, higher doses of 2,000, 3,000, and 4,000 showed body temperature change, whereas dose 5,000 caused slight discomfort. The acute toxicity studies showed that LD50 was above 5,000 mg/kg BW, as no lethality was observed/recorded in any of the groups after 7 days of treatment (Table 2). Table 2. LD50 values of the developed iscomatrix.
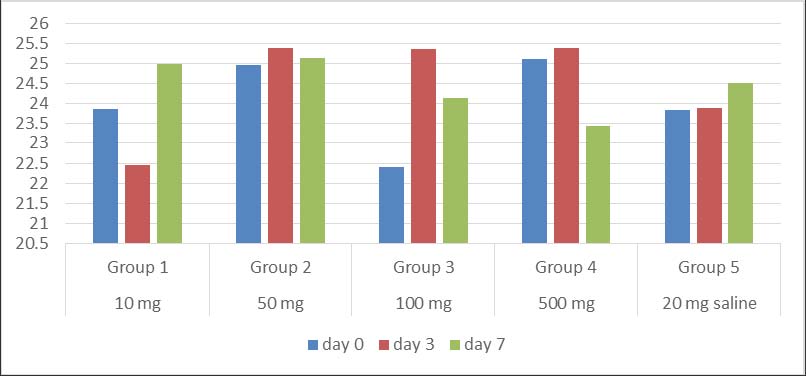
Body weight changes in the animal modelsIrrespective of the vaccine dosage, each group showed no significant (p > 0.05) weight gain/loss between days 0, 3, and 7 (Fig. 3). A general comparison of the test groups with the negative control group also showed no significant weight gain or loss (p > 0.05).
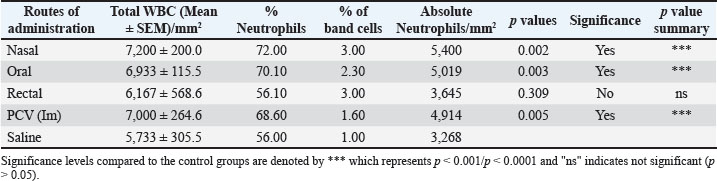
Fig. 3. Mean body weight changes after ISCOMATRIX administration. Absolute neutrophil countA notable elevation in total WBC count was observed across all animals that received the formulation at different routes apart from the saline group, reaching statistical significance (p < 0.01). The increase in the WBC resulted in some level of increase in the ANC (Table 3). Table 3. Absolute neutrophil values.
Antibody titer valuesA notable elevation (p < 0.01) in antibody A titer was recorded in all the mice immunized at different mucosal organs (Table 4). On the other hand, only intranasal immunization of the iscomatrix vaccine candidate and intramuscular immunization of the pneumococcal conjugate vaccine recorded a significant increase (p < 0.01) in antibody G titer, as presented in Table 5. Table 4. Immunoglobulin A Titer.
Table 5. Immunoglobulin G Titer.
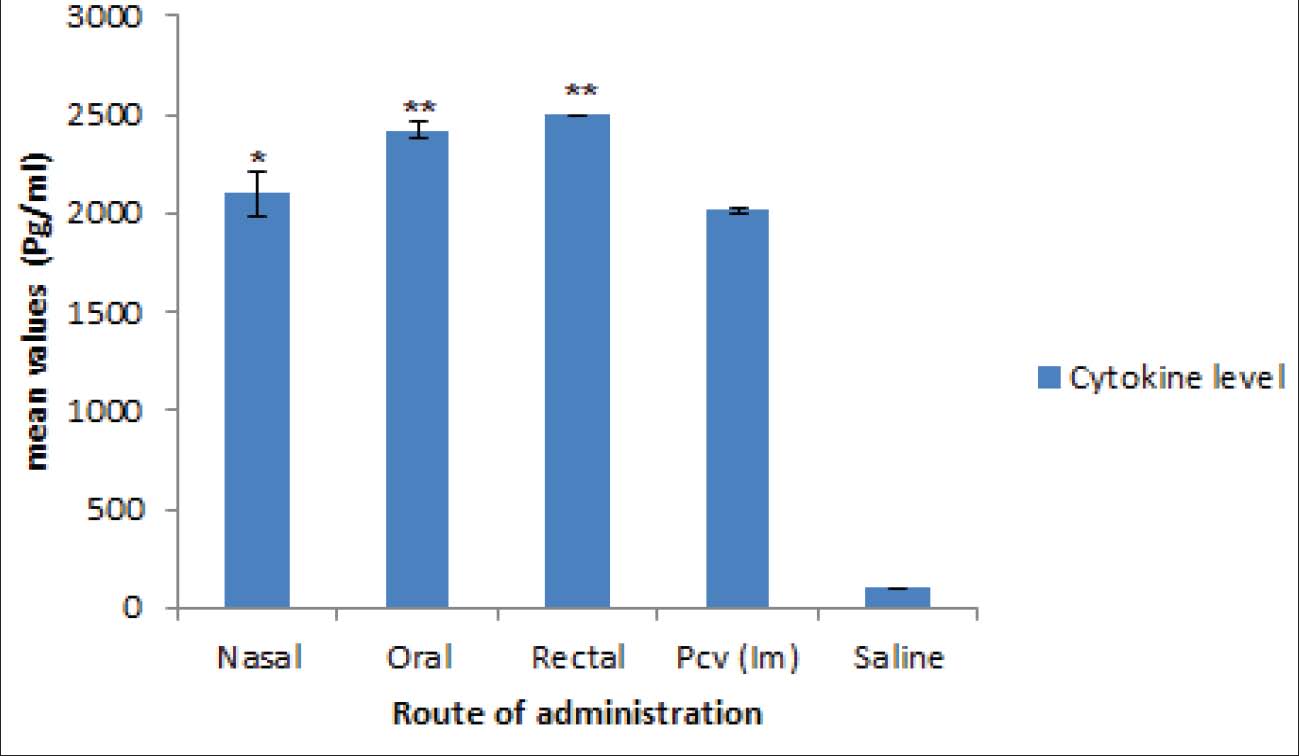
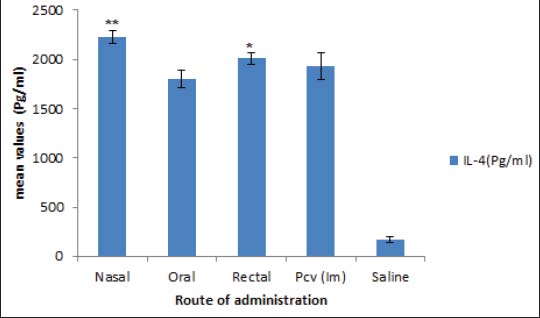
Evaluation of cellular immunityA considerable increase in the quantity of IL-4 and IFN-ᵞ indicating a corresponding elicitation (p < 0.05) of cellular immune response. As shown in Figures 4 and 5, the oral and rectal routes of administration of the vaccine candidate generated more IFN-ᵞ than the intranasal route, whereas the intranasal and rectal routes recorded more IL-4 than the oral routes.
Fig. 4. Cytokines level (IFN-ᵞ) in splenocytes. Values are presented as Mean ± SEM. Significance levels compared to the control groups are denoted by * and **, where * represents p < 0.05 and ** represents p < 0.01.
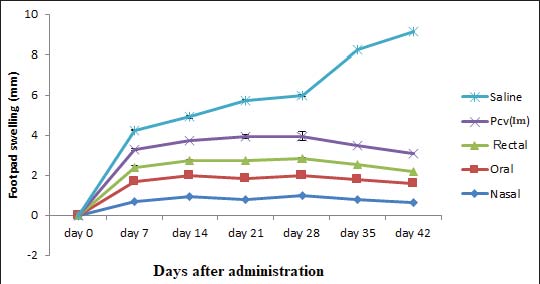
Fig. 5. Level of cytokine (IL-4) in splenocyte. The delayed hypersensitivity reactionThe progression of inflammation/swelling was rapid in all mouse groups in the first 0–7 days, with the saline group experiencing geometric progression from week 3 post-infection (Fig. 6). Some mice in the saline group died as a result of increased inflammation and swelling, whereas no progression was recorded for all other groups.
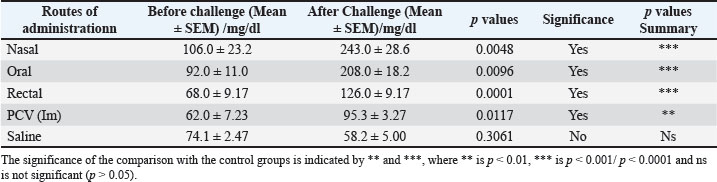
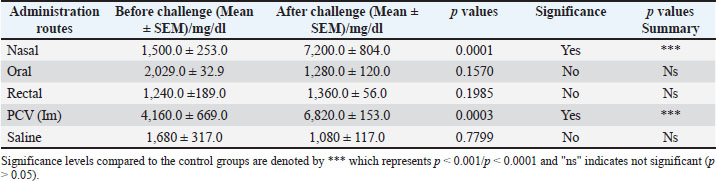
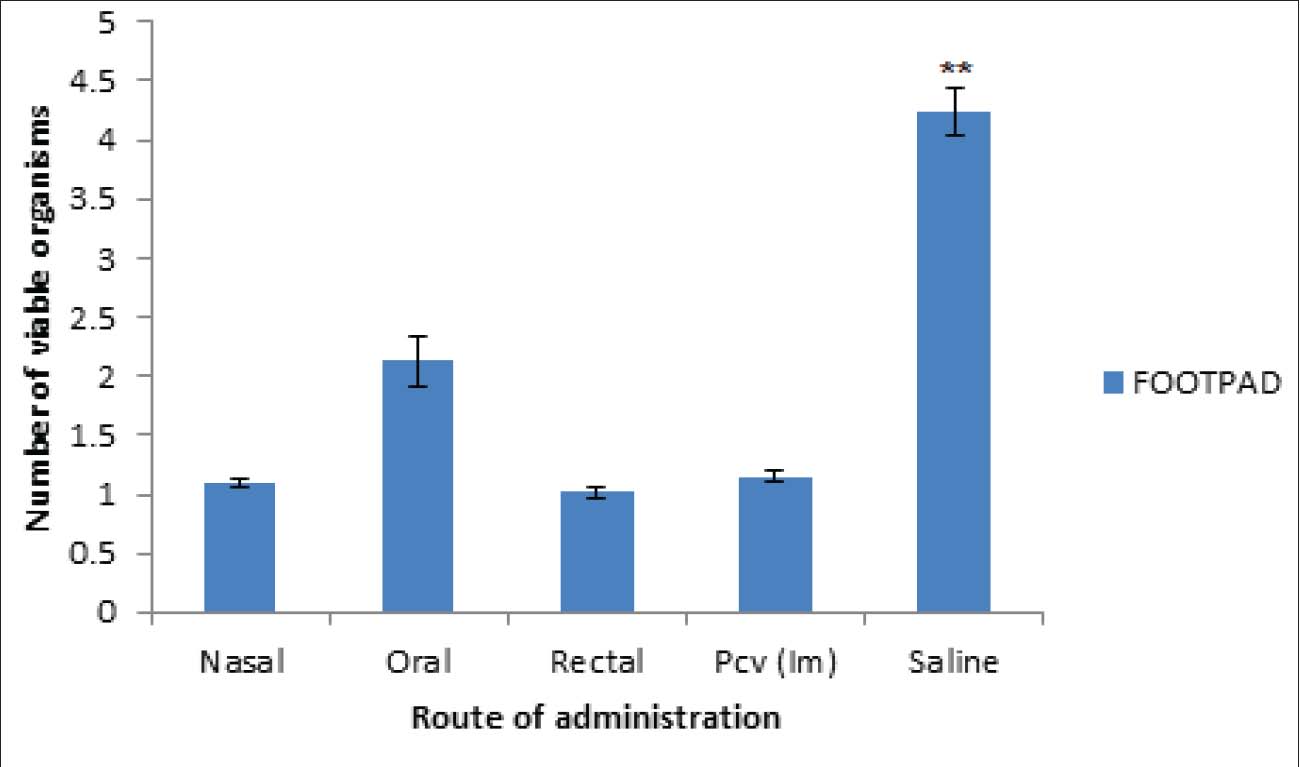
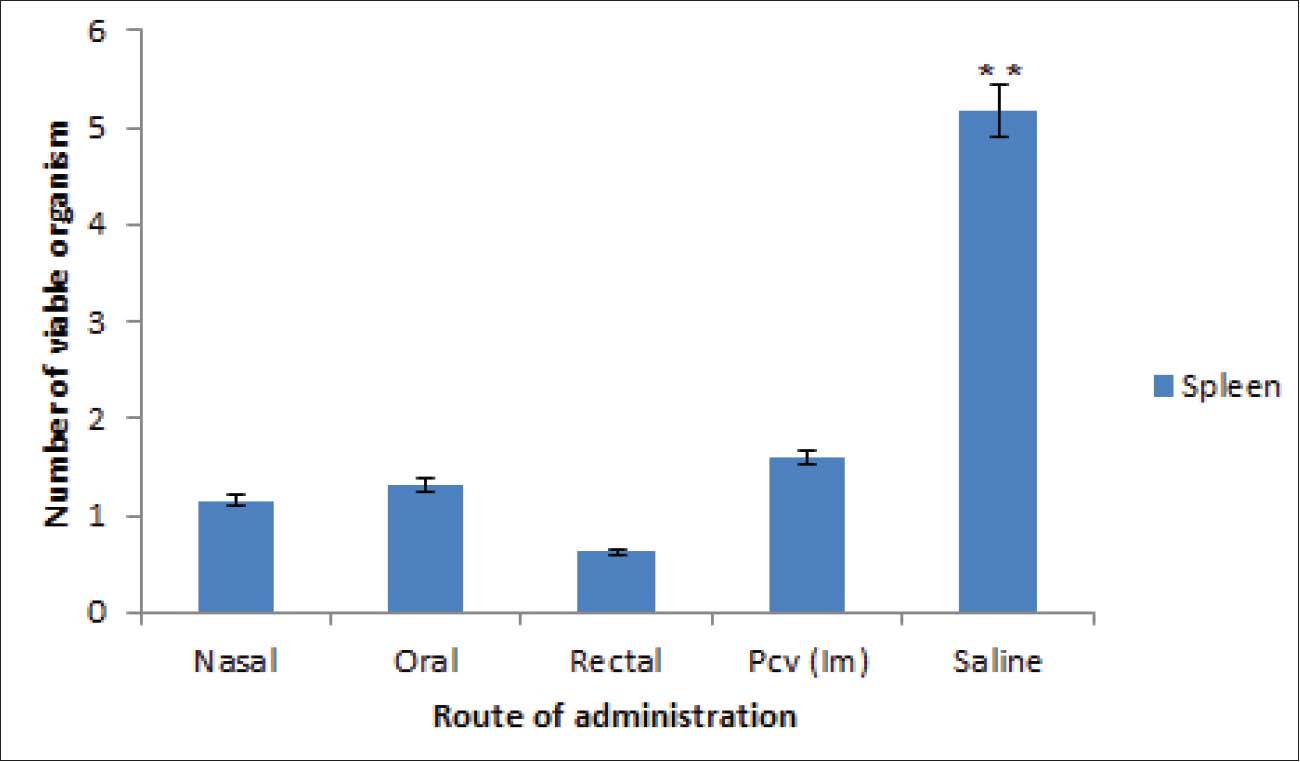
Fig. 6. Results of delayed hypersensitivity. Splenic and footpad disease burden after the challengeFigures 7 and 8 show the viable pneumococcal counts recovered from the spleen and footpad across the different experimental groups. Animals immunized via the oral, intranasal, or rectal routes demonstrated the lowest bacterial loads in the spleen, showing no significant difference from the pneumococcal conjugate vaccine (PCV) group (p > 0.05), but significantly reduced counts compared to the saline-treated controls (p < 0.05). Mice immunized intranasally or rectally also showed the least bacterial burden in the footpad; however, the differences among the immunization routes were not statistically significant when compared with the PCV control (p > 0.05). Orally immunized mice had a moderately higher bacterial count relative to the PCV group (p < 0.05), though still significantly lower than that of the saline group (p < 0.01).
Fig. 7. Number of viable bacteria in the footpad of the mice. Values are presented as Mean ± SEM. Significance levels relative to the control groups are denoted by **, where ** indicates p < 0.01.
Fig. 8. Number of viable bacteria in the spleen of mice. DiscussionThe ISCOMATRIX adjuvant comprises saponin, cholesterol, and phospholipids, which assemble into a cage-like nanoparticle structure with a diameter of approximately 40–50 nm and a surface charge of −20 mV or lower. This architecture facilitates efficient antigen delivery and presentation while also conferring strong immunomodulatory activity (Maraskovsky et al., 2009). Our developed iscomatrix has characteristics similar to other iscomatrix reported by Maraskovsky, et al. (2009) with a diameter of 47 nm and surface charge of −9.02mV (Maraskovsky et al., 2009), as presented in Table 1. A negative zeta potential was observed in the preparations, which can be partly ascribed to saponin content. The PDI was 0.21 and measures the heterogeneity of particle sizes in the mixture and the distribution of individual molecular masses of each particle. The solubility test of the formulation (iscomatrix) revealed that it was mildly soluble in water as a result of the interaction of cholesterol and saponin in a lipid membrane, and the formulations exhibited a near-neutral pH, averaging 7.06. The pH and solubility characteristics of the iscomatrix were based on the findings reported by Ojiako et al. (2019). Transmission electron microscopy was employed to verify the colloids’ size and structural features. The images revealed hollow, cage-like formations characteristic of the conventional ISCOMATRIX morphology. This morphology is represented in Fig. 2. To avoid degradation due to radiation and increase stability, the iscomatrix vaccine candidate was stored in dark/amber colored vials at a low temperature of −4oC. Its stability and potency were ascertained by the immunological assessment of the vaccine at 2 months apart, which gave rise to a significant immune response. This implies that at a low storage temperature of −4oC, the iscomatrix vaccine candidate maintained its potency and stability for more than 2 months. The lethality effect of the developed iscomatrix on mice (Table 2) revealed no fatalities within 24 hours following treatment. The prominent signs observed during this period included a warm body, elevated temperature (fever), and general discomfort in the 2,000–5,000 mg/kg dose groups. However, these signs were not evident in patients with 10, 100, and 1,000 mg/kg b.w. Dose groups and no further escalation were required. Fever is a natural component of the immune response and indicates that the body is actively reacting to the administered vaccine, thereby protecting against the targeted virus or bacterium. Handy (2024) showed that 24–35 of 100 infants who receive pneumococcal conjugate vaccine experience fever 0–7 days after vaccination (Handy, 2024). Therefore, the rise in temperature without serious illness observed in the animals given high doses of the iscomatrix vaccine candidate is an indication that the body’s defenses are engaging with the vaccine and tends to build immunity against the organism S. pneumoniae. With a lethality dose (LD50) exceeding 5,000 mg/kg b.w., the formulation is considered safe, as recorded by Garrido-Acosta et al. (2014). This finding also supported the findings of Sahgal, who reported that LD50 of the methanolic extract of Pleurotus was more than 5,000 mg/kg BW (Sahgal et al., 2010). The extract can be inferred to be safe for human consumption. Kuarta and Singh outlined the characteristics and impacts of an effective vaccine on the BW of the animal models under analysis. No significant (p > 0.05) weight gain/loss was observed in the weight assessment compared to the control group (Fig. 3). Thus, the developed iscomatrix vaccine candidate can be considered a good vaccine option, aligning with Kuarta (2006). The iscomatrix vaccine candidate’s effects on humoral and cellular immune responses via mucosal routes were assessed. This mode of delivery provides multiple benefits, including simple administration, eliminating the need for trained health care workers. In addition, it holds significant potential for inducing immunity at the entry portals of various pathogens, including Streptococcus pneumoniae (Davitt and Lavelle, 2015; Obaji et al., 2025). The iscomatrix formulation markedly boosted the systemic generation of neutrophils after mucosal immunizations in evaluating humoral responses. This is shown in the significant increase (p < 0.01) in the secretion of neutrophils in the serum of the mice immunized intranasally and orally with rectal routes having a non-significant secretion (p > 0.05) as shown in Table 3. Immune system cells act as phagocytes, recognizing and destroying harmful invaders. In addition, they function as cellular clean-up agents by engulfing dead cells and clearing away debris (Oli et al., 2019). A significant increase (p < 0.01) in antibody A titer was also recorded in all the mice immunized at different mucosal routes (Table 4). On the other hand, only intranasal iscomatrix immunization and pneumococcal conjugate vaccine intramuscular immunization recorded a significant increase (p < 0.01) in antibody G titer as presented in Table 5. The high IgA titer level recorded in the mucosal routes suggests that the formulated iscomatrix vaccine candidate can elicit a humoral immune response, making it a suitable adjuvant for prophylactic vaccines. The IgG titer recorded in the intranasal administration indicates its use in therapeutic vaccines. A marked IFN-γ response (p < 0.05) was detected in the splenocyte supernatants of mice immunized via the oral and rectal routes compared with other groups. Levels of IFN-γ were significantly higher (p < 0.01) in the orally, intranasally, and rectally immunized groups than in the saline control group, but no significant difference (p > 0.05) was observed when compared with the PCV group (Fig. 3). For IL-4, the greatest concentration was recorded in mice that received intranasal immunization (p < 0.05), although the increase was only modestly significant compared with the other groups (Figs. 4 and 5). However, all immunized groups demonstrated significantly elevated IL-4 levels relative to the saline-treated controls. A high level of protection against Streptococcus was observed, as can be deduced from the results of delayed hypersensitivity reactions and disease burden assessment in the footpad and spleen. The footpad thickness of all mouse groups increased rapidly over the first 0–7 days (first week) following the challenge. Inflammation size was reduced to an appreciable minimum (p < 0.05) in mice that received the iscomatrix vaccine candidate via the mucosal route. In all mice immunized by different routes, except the saline group, the inflammation size did not increase geometrically starting in week 3 after infection, as shown in Fig. 6. The number of viable bacteria (disease burden) reduced from the 1.5 × 10^8 cells inoculated to each animal to 1.0 × 108 for nasal and rectal administration in the footpad (Fig. 7), with a significant p-value less than 0.05 (p < 0.05). The splenic viable cell count indicated an immune response, as observed in the nasal (1.1 × 10^8), oral (1.3 × 10^8), and rectal (0.6 × 10^8) administration, as represented also in Figure 8. This is an indication that the formulated iscomatrix vaccine candidate can elicit cell-mediated immunity, which renders it an appropriate adjuvant for vaccines, as the effective stimulation of T-cell responses is required for protection against certain infectious diseases. The immunostimulatory effect of the iscomatrix through the high secretion of neutrophils and an increase in antibody titer caused the humoral immunity response. This immunomodulatory activity was recorded in the study by Oli et al. (2019). In their study, Ojiako et al. (2019) also recorded the significant immunostimulatory activities of Iscomatrix and its ability to elicit a high immune response (Ojiako et al., 2019). A significant cytokine production stimulated a cell-mediated response. ConclusionIn this study, the ISCOMATRIX formulation derived from P. ostreatus—demonstrated to be safe through standardized toxicity assays—exhibited promising potential as both an immunoadjuvant and a vaccine platform designed to induce strong humoral and cellular immune responses against multiple pneumococcal serotypes. This immunogenic effect was observed across multiple mucosal administration routes, including intranasal, oral, and rectal, even at comparatively low doses. The intranasal route produced the most pronounced immune activation, highlighting its efficacy in stimulating protective responses. Although the differences in overall efficacy between the ISCOMATRIX-based vaccine candidate and the conventional PCV were not statistically large, the strength and effectiveness of the immune responses—particularly the enhanced levels of IFN-γ and IL-4—suggest that mucosal delivery of the Pleurotus-based ISCOMATRIX may represent a viable alternative to parenteral immunization. These findings validate the dual role of ISCOMATRIX as both an adjuvant and an efficient mucosal delivery system capable of enhancing immunogenicity through Th1 and Th2 pathways. Collectively, the data support the hypothesis that integrating this novel ISCOMATRIX formulation into pneumococcal vaccination strategies could overcome critical limitations of current injectable vaccines, such as dependence on trained personnel, lower compliance, multi-dose regimens, risk of needle-stick injuries, and limited mucosal immunity. The immunological and safety profiles demonstrated here warrant further investigation and development of this platform for broader prophylactic and therapeutic applications in infectious disease control. Contributions to knowledgeThis study has established that saponin from P. ostreatus can be used to formulate an iscomatrix adjuvant for prophylactic and therapeutic uses as an adjuvant against the originally known Quil A. This newly formulated iscomatrix also served as a good delivery system for the pneumococcal vaccine, thus eliciting more humoral and cellular immunity than the already available pneumococcal conjugate vaccine. This study has also provided an alternative method of pneumococcal vaccine administration (mucosal) as opposed to conventional parenteral administration. Limitations of the studyFirst, the study did not evaluate protective immune responses against S. pneumoniae infection with specific antibodies because these antibodies were not readily available during the course of this research, the need for an alternative. The protective efficacy of the vaccine, with or without the adjuvant, following bacterial challenge was not assessed because the study aimed to compare the effect of the formulated iscomatrix vaccine candidate delivered mucosally with the parenteral pneumococcal conjugate vaccine already in use. This study aimed to solve the problems associated with the route of administration of pneumococcal vaccines. The immunomodulatory effect of the adjuvant alone could be further investigated. AcknowledgmentThe authors thank Mr. Emele Ufere for his assistance in extracting and isolating saponin. List of AbbreviationsDLS, dynamic light scattering; IM, Intramuscular; IL-4, Interleukin 4; IFN-γ, Interferon ᵞ; PCV, Pneumococcal Conjugate Vaccine; PD, polydispersity index; SEM, standard error of the mean; TEM, Transmission electron microscopy; WBC, white blood cells. Conflict of interestThe authors report no funding and no conflicts of interest in this work. FundingNone. Authors’ contributionsAkunna Perpetua Emeruwa and Chinemerem Paschaline Ogbu performed the experiments; Angus Nnamdi Oli conceived and designed the experiments, performed some statistical analyses, and edited the manuscript draft; and Malachy Chigozie Ugwu participated in designing the experiments. Akunna Perpetua Emeruwa and Adekunle Rowaiye wrote the manuscript draft. Data availabilityAll data are included in the manuscript. ReferencesAaberge, I.S. and Løvik, M. 1996. The antibody response after immunization with pneumococcal polysaccharide vaccine in splenectomized mice: the effect of re-immunization with pneumococcal antigens. Acta. Pathologica. Microbiologica. Et. Immunologica. Scandinavica. 104(4), 307–317; doi:10.1111/j.1699-0463.1996.tb00722 Badiee, A., Jaafari, M.R. and Khamesipour, A. 2007. Leishmania major: immune response in BALB/c mice immunized with stress-inducible protein 1 encapsulated in liposomes. Exp. Parasitol. 115(2), 127–134; doi:10.1016/j.exppara.2006.07.014 Behboudi, S., Morein, B. and Villacres-Eriksson, M. 1996. In vitro activation of antigen-presenting cells (APC) by defined composition of Quillaja saponaria Molina triterpenoids. Clin. Exp. Immunol. 105(1), 26–30; doi:10.1046/j.1365-2249.1996.d01-731.x Brito, L.A., Malyala, P. and O’Hagan, D.T. 2013. Vaccine adjuvant formulations: a pharmaceutical perspective. Semin. Immunol. 25(2), 130–145; doi:10.1016/j.smim.2013.05.007 Centers for Disease Control and Prevention. 2014. ABCs report: Streptococcus pneumoniae. https://www.cdc.gov/abcs/index.html Centers for Disease Control and Prevention. 2019. Pneumococcal disease. https://www.cdc.gov/pneumococcal/index.html Chang, S. and Miles, P. 1992. Mushroom biology: a new discipline. Mycologist 6(2), 64–65; doi:10.1016/S0269-915X(09)80118-7 Chao, Y., Marks, L.R., Pettigrew, M.M. and Hakansson, A.P. 2014. Streptococcus pneumoniae biofilm formation and dispersion during colonization and disease. Front. Cell. Infect. Microbiol. 4, 194; doi:10.3389/fcimb.2014.00194 Christopher, B. 2017. Vaccine adjuvants: methods and protocols. In Methods in molecular biology. Fox. Ed. London, UK: Springer, 1494, pp: 49–62. https://doi.org/ 10.1007/978-1-4939-6445-1_6 Davitt, C.J. and Lavelle, E.C. 2015. Delivery strategies to enhance oral vaccination against enteric infections. Adv. Drug Del. Rev. 91, 52–69; doi:10.1016/j.addr.2015.03.008 Fountaulakis, M.S., Dokianakis, S.N., Kornaros, M.E., Aggelis, G.G. and Lyberatos, G. 2002. Removal of phenolics in olive mill wastewaters using the white rot fungus Pleurotus ostreatus. Water. Res. 36(19), 4735–4744; doi:10.1016/S0043-1354(02)00186-1 García-Lafuente, A., Moro, C., Villares, A., Guillamón, E., Rostagno, M.A. and Martínez, J.A. 2011. Mushrooms as a source of anti-inflammatory agents. Am. J. Community. Psychol. 48(1–2), 125–141; doi:10.1007/s10464-011-9428-6 Garrido-Acosta, O., Meza-Toledo, S.E., Anguiano-Robledo, L., Valencia-Hernández, I. and Chamorro-Cevallos, G. 2014. Adaptation of Lorke’s method to determine and compare ED50 values: the cases of two anticonvulsant drugs. J. Pharmacol. Toxicol. Methods 70(1), 66–69; doi:10.1016/j.vascn.2014.05.002 Góral, I. and Wojciechowski, K. 2020. Surface activity and foaming properties of saponin-rich plant extracts. Adv. Colloid. Interface. Sci. 279, 102145; doi:10.1016/j.cis.2020.102145 Handy, L. 2024. Fever and vaccines. Vaccine Education Center, Children’s Hospital of Philadelphia, Philadelphia, PA. Available via https://www.chop.edu Haruka, M., Takuo, M., Masaki, O., Isao, H. and Kazunari, Y. 2010. A new method for the evaluation of vaccine safety based on comprehensive gene expression analysis. J. Biomed. Biotechnol. 7, 361841; doi:10.1155/2010/361841 Jafar, M., Ali, B., Zahra, M., Alireza, K. and Mohsen, T. 2018. Immunization against PR8 influenza virus with chitosan-coated ISCOMATRIX nanoparticles. Artificial Cells Nanomed. Biotechnol. 46(2), 587–593. Jaafari, M.R., Badiee, A., Khamesipour, A., Samiei, A., Soroush, D. and Barkhordari, F. 2007. The role of CpG ODN in the enhancement of immune response and protection in BALB/c mice immunized with recombinant major surface glycoprotein of Leishmania (rgp63) encapsulated in cationic liposome. Vaccine 25(29), 6107–6117; doi:10.1016/j.vaccine.2007.05.027 Jeffrey, B and Harborne, J.B. 1973. Phytochemical methods: a guide to modern techniques of plant analysis. London, UK: Chapman and Hall. Kensil, C.R., Patel, U., Lennick, M. and Marciani, D. 1991. Separation and characterization of saponins with adjuvant activity from Quillaja saponaria Molina cortex. J. Immunol. 146(2), 431–437. Kuarta, T. 2006. Minimum requirements for biological products. National Institute of Infectious Diseases. Lövgren Bengtsson, K., Morein, B. and Osterhaus, A.D. 2011. ISCOM technology-based Matrix-M adjuvant: success in future vaccines relies on formulation. Expert. Rev. Vaccines. 10(3), 401–403; doi:10.1586/erv.11.20 Malihe, M., Zuleeza, A., Lai, T.G. and Chit, L.P 2020. Development of next generation Streptococcus pneumoniae vaccines conferring broad protection. Vaccines 8, 132. Maraskovsky, E., Schnurr, M., Wilson, N.S., Robson, N.C., Boyle, J. and Drane, D. 2009. Development of prophylactic and therapeutic vaccines using the ISCOMATRIX adjuvant. Immunol. Cell Biol. 87(5), 371–376; doi:10.1038/icb.2009.25 Musher, D.M. and Thorner, A.R. 2014. Community-acquired pneumonia. N. Engl. J. Med. 371(17), 1619–1628; doi:10.1056/NEJMra1312885 Negi, J., Pant, G.J., Singh, P. and Rawat, M.S. 2011. High-performance liquid chromatography analysis of plant saponins: an update 2005–2010. Pharmacognosy Rev. 5(10), 155–158; doi:10.4103/0973-7847.91109 Ngai, P.H.K. and Ng, T.B. 2004. A ribonuclease with antimicrobial, antimitogenic and antiproliferative activities from the edible mushroom Pleurotus sajor-caju. Peptides 25(1), 11–17; doi:10.1016/j.peptides.2003.12.005 Obaji, M., Oli, A. and Ugwu, M. 2025. Characterization of Streptococcus pneumoniae isolated from paediatric patients in a tertiary hospital in Anambra State (Unpublished doctoral dissertation). Nnamdi Azikiwe University, Awka, Anambra State. Obaji, M.U., Emeruwa, A.P., Oli, A.N. and Ugwu, M.C. 2025. Innovative approaches in pneumococcal vaccine development: harnessing natural compounds and advanced delivery systems. J. Curr. Biomed. Res. 5(1), 1947–1970; https://doi.org/10.54117/ Ojiako, C.M., Okoye, E.I., Oli, A.N., Ike, C.J., Esimone, C.O. and Attama, A.A. 2019. Preliminary studies on formulation of immune stimulating complexes using saponin from Carica papaya leaves. Heliyon 5(5), 1897; doi:10.1016/j.heliyon.2019.e01897 Oli, A.N., Emeruwa, A.P., Ekwunife, O.I., Okoye, E.L., Anyaoku, C.S., Ibeanu, G.C., Obi, E., Ihekwereme, C.P. and Obaji, M. 2019. Pleurotus ostreatus extract enhances the phagocytic actions of neutrophils against Streptococcus pneumoniae. Ann. Med. Health. Sci. Res. 9(3), 499–508. Oli, A.N., Uchenna, A.R., Oli, U.C. and Agu, R.U. 2015. Safety evaluation in mice of the childhood immunization vaccines from two South-eastern states of Nigeria. Asian Pac. J. Trop. Biomed. 5(2), 132–137; doi:10.1016/S2221-1691(15)30162-9 Sahgal, G., Ramanathan, S., Sasidharan, S., Mordi, M.N., Ismail, S. and Mansor, S.M. 2010. Brine shrimp lethality and acute oral toxicity studies on Swietenia mahagoni (Linn.) Jacq. seed methanolic extract. Pharmacognosy Res. 2(4), 215–220; doi:10.4103/0974-8490.69105 Rowaiye, A., Wilfred, O.I., Onuh, O.A., Bur, D., Oni, S., Nwonu, E.J., Ibeanu, G., Oli, A.N. and Wood, T.T. 2022. Modulatory effects of mushrooms on the inflammatory signaling pathways and pro-inflammatory mediators. Clin. Complement. Med. Pharmacol. 2(4), 100037; doi:10.1016/j.ccmp.2022.100037 Sasidharan, S., Mordi, M.N., Ismail, S., Mansor, S., Sahgal, G. and Ramanathan, S. 2010. Brine shrimp lethality and acute oral toxicity studies on Swietenia mahagoni (Linn.) Jacq. seed methanolic extract. Pharmacognosy Res. 2(4), 215–220; doi:10.4103/0974-8490.69105 Su, F., Patel, G.B., Hu, S. and Chen, W. 2016. Induction of mucosal immunity through systemic immunization: phantom or reality?. Hum. Vaccines. Immunotherapeutics. 12(4), 1070–1079; doi:10.1080/21645515.2015.1114198 Sun, H.X., Xie, Y. and Ye, Y.P. 2009. ISCOMs and ISCOMATRIX. Vaccine 27(33), 4388–4401; doi:10.1016/j.vaccine.2009.05.041 Taswell, C. 1981. Limiting dilution assays for determination of immunocompetent cell frequencies. I. Data analysis. J. Immunol. 126(4), 1614–1619. Titus, R.G., Marchand, M., Boon, T. and Louis, J.A. 1985. A limiting dilution assay for quantifying Leishmania major in tissues of infected mice. Parasite. Immunol. 7(5), 545–555; doi:10.1111/j.1365-3024.1985.tb00098.x Wang, H.X. and Ng, T.B. 2000. Isolation of a novel ubiquitin-like protein from Pleurotus ostreatus mushroom with anti-human immunodeficiency virus, translation-inhibitory and ribonuclease activities. Biochem. Biophys. Res. Commun. 276(2), 587–593; doi:10.1006/bbrc.2000.3480 World Health Organization. 2019. SAGE working group on pneumococcal conjugate vaccines. Available via https://www.who.int/immunization/policy/sage/sage_wg_pcv_dec2016/n// | ||


| How to Cite this Article |
| Pubmed Style Emeruwa AP, Oli AN, Ugwu MC, Rowaiye AB, Ogbu CP. Mucosal delivery of pleurotus-based ISCOMATRIX elicits humoral and cellular immunity against pneumococcal infection. Open Vet. J.. 2026; 16(5): 3203-3217. doi:10.5455/OVJ.2026.v16.i5.60 Web Style Emeruwa AP, Oli AN, Ugwu MC, Rowaiye AB, Ogbu CP. Mucosal delivery of pleurotus-based ISCOMATRIX elicits humoral and cellular immunity against pneumococcal infection. https://www.openveterinaryjournal.com/?mno=289028 [Access: June 26, 2026]. doi:10.5455/OVJ.2026.v16.i5.60 AMA (American Medical Association) Style Emeruwa AP, Oli AN, Ugwu MC, Rowaiye AB, Ogbu CP. Mucosal delivery of pleurotus-based ISCOMATRIX elicits humoral and cellular immunity against pneumococcal infection. Open Vet. J.. 2026; 16(5): 3203-3217. doi:10.5455/OVJ.2026.v16.i5.60 Vancouver/ICMJE Style Emeruwa AP, Oli AN, Ugwu MC, Rowaiye AB, Ogbu CP. Mucosal delivery of pleurotus-based ISCOMATRIX elicits humoral and cellular immunity against pneumococcal infection. Open Vet. J.. (2026), [cited June 26, 2026]; 16(5): 3203-3217. doi:10.5455/OVJ.2026.v16.i5.60 Harvard Style Emeruwa, A. P., Oli, . A. N., Ugwu, . M. C., Rowaiye, . A. B. & Ogbu, . C. P. (2026) Mucosal delivery of pleurotus-based ISCOMATRIX elicits humoral and cellular immunity against pneumococcal infection. Open Vet. J., 16 (5), 3203-3217. doi:10.5455/OVJ.2026.v16.i5.60 Turabian Style Emeruwa, Akunna Perpetua, Angus Nnamdi Oli, Malachy Chigozie Ugwu, Adekunle B. Rowaiye, and Chinemerem Paschaline Ogbu. 2026. Mucosal delivery of pleurotus-based ISCOMATRIX elicits humoral and cellular immunity against pneumococcal infection. Open Veterinary Journal, 16 (5), 3203-3217. doi:10.5455/OVJ.2026.v16.i5.60 Chicago Style Emeruwa, Akunna Perpetua, Angus Nnamdi Oli, Malachy Chigozie Ugwu, Adekunle B. Rowaiye, and Chinemerem Paschaline Ogbu. "Mucosal delivery of pleurotus-based ISCOMATRIX elicits humoral and cellular immunity against pneumococcal infection." Open Veterinary Journal 16 (2026), 3203-3217. doi:10.5455/OVJ.2026.v16.i5.60 MLA (The Modern Language Association) Style Emeruwa, Akunna Perpetua, Angus Nnamdi Oli, Malachy Chigozie Ugwu, Adekunle B. Rowaiye, and Chinemerem Paschaline Ogbu. "Mucosal delivery of pleurotus-based ISCOMATRIX elicits humoral and cellular immunity against pneumococcal infection." Open Veterinary Journal 16.5 (2026), 3203-3217. Print. doi:10.5455/OVJ.2026.v16.i5.60 APA (American Psychological Association) Style Emeruwa, A. P., Oli, . A. N., Ugwu, . M. C., Rowaiye, . A. B. & Ogbu, . C. P. (2026) Mucosal delivery of pleurotus-based ISCOMATRIX elicits humoral and cellular immunity against pneumococcal infection. Open Veterinary Journal, 16 (5), 3203-3217. doi:10.5455/OVJ.2026.v16.i5.60 |

