Open Veterinary Journal, (2026), Vol. 16(5): 3237-3246
Short Communication
10.5455/OVJ.2026.v16.i5.63
Effects of the methanolic extract of Tradescantia spathacea on nitro-oxidative and chemokine responses in peritoneal macrophages from BALB/c and C57BL/6 mice
Diana L. Chávez-Aviña1, Ángeles G. Lugo-Díaz1, Kibsaim Franco-Villanueva2,
Víctor I. Lozano-Vielmas2, Gustavo Hernández-Vidal2, Raymundo A. Pérez-Hernández1,
Adolfo Soto-Dominguez3 and Uziel Castillo-Velázquez2*
1Faculty of Biological Sciences, Universidad Autónoma de Nuevo León (UANL), San Nicolás de los Garza, Mexico
2Faculty of Veterinary Medicine and Animal Science, Universidad Autónoma de Nuevo León (UANL), Escobedo, Mexico
3Faculty of Medicine, Universidad Autónoma de Nuevo León (UANL), Mitras Centro Monterrey, Mexico
*Corresponding Author: Uziel Castillo-Velázquez. Faculty of Veterinary Medicine and Animal Science, Universidad Autónoma de Nuevo León (UANL), Escobedo, Mexico. Email: uziel.castillovl [at] uanl.edu.mx
Submitted: 03/09/2025 Revised: 27/03/2026 Accepted: 07/04/2026 Published: 31/05/2026
© 2025 Open Veterinary Journal
This is an Open Access article distributed under the terms of the Creative Commons Attribution-Non Commercial-No Derivatives License (http://creativecommons.org/licenses/by-nc-nd/4.0/), which permits non-commercial re-use, distribution, and reproduction in any medium, provided the original work is properly cited, and is not altered, transformed, or built upon in any way.
ABSTRACT
Background: During innate immunity, peritoneal macrophages (PMs) orchestrate early nitro-oxidative and chemokine responses, and their polarization is influenced by mouse strain-dependent metabolic programs. Tradescantia spathacea is a medicinal plant with reported anti-inflammatory and antioxidant properties; however, its effects on macrophage nitro-oxidative and chemokine responses remain unclear.
Aim: To investigate the effects of a methanolic extract of T. spathacea (MET.s) on nitro-oxidative and chemokine responses in PMs from BALB/c and C57BL/6 mice.
Methods: Lipopolysaccharide (LPS)-elicited PMs were obtained from female BALB/c and C57BL/6 mice. The cytotoxicity and half-maximal inhibitory concentrations (IC50) of MET.s were determined by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide assay. Nitric oxide (NO) production was quantified using the Griess reaction. Gene expression of iNOS, ARGII, TGF-β, MCP, and MIP was evaluated by real-time PCR in PMs stimulated with IL-4 or LPS, alone or in combination with strain-specific IC50 concentrations of MET.s.
Results: MET.s did not reduce mitochondrial activity in either strain; IC50 values were 4.76 µg/ml for BALB/c and 81.92 µg/ml for C57BL/6 PMs. BALB/c PMs exhibited a modest but significant increase in NO at 50 µg/ml, whereas C57BL/6 PMs exhibited decreased NO at the same concentration, inverting the expected strain pattern. In BALB/c PMs, MET.s alone or in combination with LPS or IL-4 favored iNOS and TGF-β expression, suggesting a pro-nitro-oxidative yet regulated profile. In C57BL/6 PMs, LPS+MET.s downregulated iNOS while upregulating ARGII and TGF-β, while MET.s selectively increased MIP with concurrent MCP reduction in both LPS- and IL-4-driven conditions, indicating a shift towards a balanced chemokine-mediated recruitment.
Conclusion: The MET.s exerts non-cytotoxic, strain-dependent immunomodulatory effects on PMs, reshaping nitro-oxidative output and chemokine gene expression to favor a context-dependent balance between activation and regulation. These findings support its potential as a natural immunomodulator and justify further mechanistic and in vivo studies.
Keywords: BALB/c, C57BL/6, Chemokines, Peritoneal macrophages, Tradescantia spathacea.
Introduction
A regulatory process on innate immune response is fundamental for maintaining tissue homeostasis and preserving the organism’s integrity when facing possible pathogenic agents. In this context, macrophages emerge as key effector cells in the induction and resolution of inflammatory processes (Ross et al., 2021). Their remarkable versatility enables them to play dual roles during the immune response. At a basal state, peritoneal macrophages (PMs) exhibit a naïve phenotype, which can be modified in response to microenvironmental cues, leading to two main activation phenotypes: classically activated (M1) and alternatively activated (M2) macrophages (Breda et al., 2022).
Pro-inflammatory cytokines secreted by Th1 lymphocytes, including interferon-gamma and tumor necrosis factor-alpha (TNF-α), induce M1macrophage polarization. These macrophages exhibit strong microbicidal activity and secrete inflammatory mediators. In contrast, anti-inflammatory cytokines secreted by Th2 lymphocytes, such as IL-4 and IL-13, activate M2 macrophages and exhibit functions associated with tissue repair and homeostasis (Restrepo et al., 2025).
Murine models such as C57BL/6 and BALB/c present a Th1- and Th2-biased immune response, respectively (Restrepo et al., 2025), and differ in amino acid metabolism, particularly arginine (Øynebråten et al., 2019). It has been established that the metabolic environment plays a critical role in determining macrophage polarization; for instance, elevated glucose metabolism and glycolysis are hallmarks of M1 macrophages, while M2 macrophages rely on fatty acid oxidation and enhanced oxidative phosphorylation (OXPHOS) (Han et al., 2025). Available data indicate that macrophages derived from each strain exhibit distinct activation pathways in response to the same stimuli, such as lipopolysaccharide (LPS), steering a characteristic response of M1 and M2 macrophages. In C57BL/6 mice, nitric oxide synthase 2 (NOS2) expression leads to elevated levels of nitric oxide (NO) from L-arginine, favoring differentiation to an M1 pro-inflammatory phenotype. The production of reactive nitrogen species inhibits OXPHOS, driving a metabolic shift to glycolysis for adenosine triphosphate production (Mamani-Huanca et al., 2021). On the other hand, BALB/c macrophages metabolize arginine via arginase-1, promoting ornithine and urea production, which induces M2 macrophage polarization (Øynebråten et al., 2019).
The peritoneal cavity harbors heterogeneous macrophage subsets, including long-lived tissue-resident large peritoneal macrophages (LPMs) and recruited inflammatory monocyte-derived macrophages, among tissue-resident cell populations under eliciting conditions (Arabpour et al., 2021). In the present study, macrophages were collected after a brief intraperitoneal exposure to LPS prior to lavage; therefore, the resulting cultures are more accurately described as LPS-elicited PMs (peritoneal exudate macrophages), rather than exclusively resident LPMs. Resident LPMs arise during embryonic development, are long-lived, and remain functionally confined to the peritoneal cavity, where they contribute to homeostasis (Ardavín et al., 2023). However, elicitation protocols may increase the proportion of recruited cells, which should be considered when interpreting polarization-related readouts. Despite this, PMs remain a highly accessible and reactive system to investigate immunomodulation under standardized inflammatory baseline conditions.
In the last couple of years, the need for new natural immunomodulatory agents has gained particular interest. Mexico is known worldwide as the second country with an ancestral tradition in the use of medicinal plants due to its wide range of environmental conditions and the diversity of species found within its territory (Lucía et al., 2021). Tradescantia spathacea Sw., commonly known as “purple maguey,” a Commelinaceae species native to the tropical regions of America (Tan and Kwan, 2020), contains several bioactive compounds with a total phenol content of 203.9 ± 16.3 mg GAE/100 g sample, a total tannin content of 20.6 ± 2.3 mg TAE/100 g sample, and a total flavonoid content of 10.8 ± 2.9 mg RE/100 g sample (Tan et al., 2014). Notably, the flavonol kaempfero 1 and the phenol bracteanolide A13 inhibit protein tyrosine phosphate 1B, a key negative regulator of insulin and leptin signaling pathways that also modulates immune cell activation (Kushwaha et al., 2024).
While preliminary studies have shown that T. spathacea extracts promote human lymphocyte proliferation (Sriwanthana et al., 2007) and modulate astrocyte-mediated neuroinflammation (Lopes et al., 2024), its immunomodulatory effects remain unexplored. Tu et al. (2019)performed a similar assay and demonstrated that isolates and synthesized derivatives from T. albiflora exhibited enhanced NO inhibitory activity on LPS-stimulated RAW 264.7 cells. However, no studies have analyzed the effects of T. spathacea on PMs or whether these effects differ between Th1- and Th2-biased metabolism.
Therefore, the present study aimed to evaluate the immunomodulatory effects of T. spathacea on strain-specific immune responses to elucidate the immunological properties of the plant and its potential interactions with the host immune system.
Materials and Methods
Collection of plant material
The plant material was collected from a domestic garden located in Yajalón, Chiapas, México (17.165745, -92.330700), on January 19th, 2024. Only T. spathacea leaves exceeding 25 cm in length were taken. These leaves were cleaned with a damp cloth and packed for transportation to Universidad Autónoma de Nuevo León. They were then weighted (approximately 500 g), cut into squares, and dried in a drying oven at 37.7°C for 72 hours, resulting in a dry weight of 47.4 g.
Methanolic extract preparation
The dried leaves were placed in two Erlenmeyer flasks containing 500 ml of methanol and left to macerate for 1 week at room temperature under constant agitation. The plant material was then separated using filter paper. The extract was filtered through a filter paper, and the solvent was removed by reduced pressure using a rotary evaporator at a bath temperature of 40°C and an agitation speed of 80–120 rpm.
The resulting concentrated solution was stored in a sterile flask and desiccated at 37.7°C for 48 hours to remove any remaining methanol. Once dried, the total content of Tradescantia spathacea methanolic extract (MET.s) was weighed to determine the final plant yield. The dried extract was stored at 4°C until further use.
A partial phytochemical characterization of the methanolic extract was subsequently performed using colorimetric assays, including Salkowski, Molisch, Dragendorff, and phenolic compound tests, following standard qualitative procedures to detect the presence of major groups of secondary metabolites.
Maintaining and storage of animal models
Three female mice aged 10 weeks and an average weight of 35 g were kept with food and water ad libitum, under 12 hours light/dark cycles, at an ambient temperature of 22°C ± 1°C. Mice were housed in acrylic cages (29 × 22 × 14 cm) with stainless-steel wire lids (Birdcage, Birds Park, USA) equipped with a 60 ml glass water bottle (Redkite, Mexico) and fed with commercial food (FormuLab Chows 5008; Purina, EUA). They were randomly arranged into two groups. Animal handling was performed according to the Mexican Official Standard for the production, care, and use of laboratory animals (NOM-062-ZOO-1999) and the Bioethics and Animal Welfare Committee of the Faculty of Veterinary Medicine, Universidad Autónoma de Nuevo León guidelines.
Extraction of PMs
Each mouse was anesthetized intramuscularly with a dose of 7.5 mg/kg of a dissociative agent (Zoletil® 50, Virbac). Once under deep anesthesia, they were sacrificed by cervical dislocation following the Mexican Standard (NOM-062-ZOO-1999). PMs from three independent animals per strain (n=3 biological replicates per condition) were used in all experiments, and each condition was assayed in triplicate. This sample size is commonly employed in in vitro macrophage studies and, according to a priori power estimates assuming a large effect size (Cohen’s d ≥ 1.3), provides >80% power at α=0.05 to detect ≥1.5-fold differences between treatments.
Macrophage activation was initially induced by injecting 1 ml of Dulbecco’s Modified Eagle Medium (DMEM) supplemented with 10% fetal bovine serum (FBS) + 100 ng/ml LPS into the peritoneal cavity of BALB/c and C57BL/6 mice and allowed to rest for 2 hours. This brief intraperitoneal LPS exposure was used as an elicitation/priming step prior to lavage to standardize an inflammatory baseline and obtain sufficient cell yield; accordingly, we refer to these cultures as LPS-elicited PMs (peritoneal exudate macrophages) throughout the manuscript. Macrophages were then isolated by peritoneal lavage using 10 ml of DMEM medium 10% FBS per mouse. Half of the total volume was injected into the peritoneum, and the abdominal wall was then gently massaged for 10–25 seconds to detach adherent macrophages, as reported by De De Jesus et al. (2022).
Lavage was performed until the maximum possible amount of medium (approximately 8 ml) was recovered. The recovered medium was centrifuged at 1,750 rpm for 5–10 minutes and the resulting cell pellet was resuspended in 1 ml DMEM 10% FBS and manually counted using a Neubauer chamber by mixing 10 µl of the cell suspension with 10 µl of Trypan Blue (Sigma-Aldrich). Finally, the cells were seeded in two T-25 culture flasks and incubated at 37°C with 5% CO2 for 24 hours.
Cytotoxicity assay
The 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) assay was performed to determine the cellular metabolic activity. A 96-well plate was assembled with the equivalent of 10,000 cells/well based on the manually determined cell count obtained from Trypan Blue assay, corresponding to BALB/c and C57BL/6 PMs, for subsequent incubation at previously described conditions.
The culture medium was then removed and replaced with the corresponding treatment for each experimental group. A positive control [LPS (100 ng/ml)], a negative control (DMEM 10% FBS), and six different treatments were administered in independent triplicates. Except for controls, culture medium was replaced with 100 µl of DMEM + MET.s at concentrations ranging from 200 to 6.25 µg/ml in serial 1:2 dilutions in all the remaining wells. The samples were incubated as previously described.
After incubation, each treatment was removed and replaced with 100 µl of 0.25 mg/ml MTT (Sigma-Aldrich) and incubated for 3 hours under the same conditions. MTT was then removed, and 100 µl of dimethyl sulfoxide (Sigma-Aldrich) were added to dissolve formazan crystals. The cell plate was incubated and protected from light for 30 minutes.
Finally, the cell plate was analyzed using an ELISA Epoch Reader (Epoch 2 Microplate Spectrophotometer (BioTek Instruments, Inc., Winooski, VT) at a wavelength of 540–570 nm.
Data were analyzed using the Quest Graph™ inhibitory concentrations (IC50) Calculator based on dose-response curves (AAT Bioquest Inc., Pleasanton, CA).
NO production
The induced production of NO in murine macrophages by MET.s was determined using Griess reagent. A reference curve was initially created using a 100 µM nitrite solution and 6 serial dilutions (100, 50, 25, 12.5, 6.25, 3.13, 1.56, and 0 μM) with DMEM medium in a 96-well microplate.
To obtain the experimental samples, PM cultures were stimulated with different concentrations of MET.s (from 200 to 6.25 µg/ml in serial 1:2 dilutions). After incubation at 37°C for 24 hours, the supernatant was recovered for testing.
50 µl of the sample (culture supernatant or nitrite solution for the reference curve) and 50 µl of sulfanilamide at room temperature were added to each well and incubated for 5–10 minutes, protected from light. 50 µl of N-(1-Naphthyl)ethylenediamine dihydrochloride solutions were then added and incubated for an additional 10 minutes. Readings were performed at 540 nm within the following 30 minutes.
PMs stimulation
The murine PM isolation process was performed until six culture flasks were obtained for each strain. They were incubated under the same conditions previously described until use.
Each culture flask was labeled with one of the following experimental groups (Table 1), adding the IC50 of the MET.s corresponding to each murine strain when required by the experimental group:
Table 1. Experimental groups applied to each strain.

Since cells were adherent, the culture medium contained in each flask was removed and replaced with 5 ml of the corresponding treatment, excluding the negative control. Cell culture flasks were maintained under incubation conditions for 24 hours.
RNA extraction by single-step method
Total RNA was extracted following the one-step method described by Chomczynski (1993) using the RNAzol® reagent (guanidinium thiocyanate; Molecular Research Center, Cincinnati, OH). Lysates were homogenized and processed with chloroform (Sigma-Aldrich), followed by isopropanol (Sigma-Aldrich) precipitation and ethanol washes. The recovered pellet was washed with 200 µl of 75% ethanol and centrifuged at 7,500 × g for 15 minutes at 4°C. This step was repeated twice.
After the final wash, the supernatant was carefully removed, and the recovered RNA pellet was air-dried at room temperature before being resuspended in 20 µl of nuclease-free water. Samples were incubated at 60°C for 10–15 minutes in a thermoplate, and RNA quantification was performed using an Elisa Epoch Reader (Take3 Microplate Spectrophotometer (BioTek Instruments, Inc., Winooski, VT), loading 2 µl per well and using nuclease-free water as the blank.
Complementary DNA (cDNA) synthesis
cDNA synthesis was performed using the commercial reverse (RV) transcription kit ImProm-II™ Reverse Transcription System (Promega®, USA). For each reaction, a mix containing 4 μl of MgCl2 (25 mM), 1 μl of Oligo (dT)15, 1 μl of ImPromII ™ reverse transcriptase, 1,500 ng of RNA template, and nuclease-free water to a final volume of 20 µl was prepared. Reactions were performed on 200 µl Eppendorf® tubes.
Samples were incubated in a Veritti® thermal cycler (Applied Biosystem®, USA) and quantified by spectrophotometry (EPOCH, BioTek, Canada), and the resulting products were stored at −20°C until use.
Real-time PCR (qPCR) for cytokine gene expression evaluation
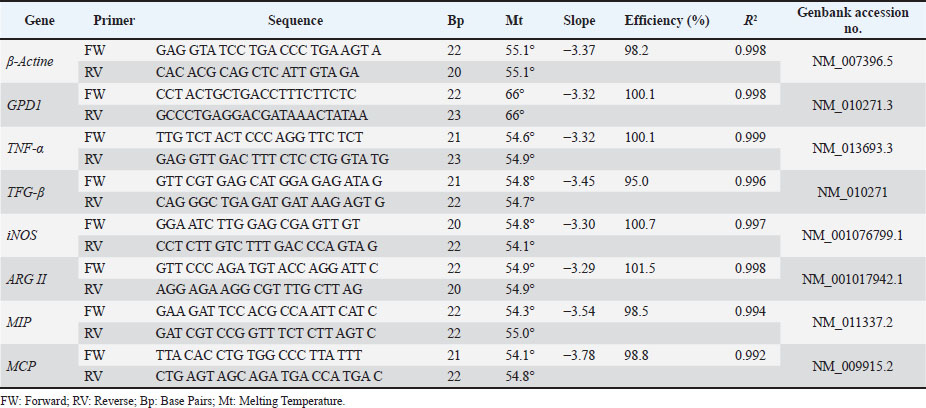
Relative quantification of gene sequences was performed by qPCR. Gene expression was determined using the ΔΔCt method, with β-actin as the reference gene for normalization of the genes of interest: TNF-α, TGF-β, iNOS, ARG II, MCP, and MIP. In accordance with Minimum Information for Publication of Quantitative Real-Time PCR Experiments (MIQE) guidance, reference gene suitability was assessed by monitoring β-actin (Actb) Ct variation across strains and treatments and by cross-validating normalization using GPD1 as an alternative reference gene; because this was a pilot short communication, only Actb-based normalization is reported in the main text for brevity. Table S1 lists the forward (FW) and RV primer sequences used. Standard curves for all target and reference genes showed linearity (R²=0.992–0.999) and amplification efficiencies between 95% and 102%, indicating optimal primer performance according to MIQE guidelines.
A 96-well plate was prepared for each mouse strain. Each qPCR reaction had a final volume of 20 µl, consisting of 5 µl of extracted cDNA (or nuclease-free water for the no-template control, NTC) and 15 µl of master mix. The master mix (15 µl per reaction) contained 10 µl of GoTaq, 4 µl of nuclease-free water, 0.5 µl of FW primer, and 0.5 µl of RV primer. Then, 15 µl of the master mix was dispensed into each well, and 5 µl of the corresponding template was added.
qPCR conditions were as follows: an initial step at 50°C for 2 minutes, followed by polymerase activation at 95°C for 2 minutes, and 40 amplification cycles consisting of denaturation at 95°C for 30 seconds, annealing at 55°C for 30 seconds, and extension at 72°C for 45 seconds. A melting curve was generated by increasing the temperature from 60°C to 95°C. NTC reactions were performed on each plate, and all samples were run in triplicate.
Statistical analysis
Data were analyzed using GraphPad Prism 8 (GraphPad Software, Inc., La Jolla, CA). Statistically significant differences between groups were analyzed using a two-way ANOVA followed by Tukey’s multiple comparison test. Results are expressed as mean ± standard deviation of the mean of three replicates with three independent assays. A 95% confidence level was applied, and differences were considered statistically significant at p < 0.05.
Ethical approval
This study was conducted at the Faculty of Veterinary Medicine and Zootechnics at the Universidad Autónoma de Nuevo León, General Mariano Escobedo, Nuevo León, Mexico. Bioethics and Animal Welfare Committee of the Faculty of Veterinary Medicine and Zootechnics at the Universidad Autónoma de Nuevo León (FMVZ/UANL) study number: 063/2023 dictamen 073/202.
Results and Discussion
Methanolic extract yield
From 47.7 g of dry T. spathacea leaf material, a total of 1.1 g of methanolic extract was obtained after the rotoevaporation process, corresponding to a yield of 2.3% (w/w), as calculated using the following formula:

Additionally, the solvent-to-biomass ratio was determined to be 10.5 ml/g of dry biomass, which was calculated according to the following equation:

The selection of methanol as the extraction solvent was based on its efficiency in isolating bioactive compounds from plant materials. Compared to other organic solvents, methanol provides higher extraction yields and greater extract purity. Its polarity allows for the efficient solubilization of a wide range of phytochemicals, including phenolic compounds, flavonoids, and other metabolites with antioxidant and antimicrobial potential, supporting its suitability for phytochemical and bioactivity analyses (Osmić et al., 2018).
The partial phytochemical characterization of the methanolic extract revealed the presence of lactones, coumarins, saponins, flavonoids, and alkaloids, as well as the possible presence of steroids and terpenes. These results suggest that T. spathacea leaves contain a wide variety of secondary metabolites with potential biological activity. The abundance of steroids and terpenes is particularly interesting, as these compounds are associated with anti-inflammatory, antimicrobial, and antioxidant properties in other Tradescantia species (Butnariu et al., 2022).
Evaluation of mitochondrial activity
Cellular metabolic activity was assessed using the MTT assay to evaluate the cytotoxicity of MET.s. PMs from both mouse strains were exposed to a concentration range of MET.s (200, 100, 50, 25, 12.5, and 6.25 µg/ml), including negative and positive controls. In BALB/c macrophages, MET.s did not reduce mitochondrial activity relative to the negative control (normalized to 100%); values remained comparable across the tested concentrations, with only a slight, non-significant increase at some doses. Similarly, in C57BL/6 macrophages, MET.s did not induce significant changes in mitochondrial activity compared with the negative control. The IC50 values were estimated from the concentration-response curve derived from the MTT concentration series (Fig. 1), yielding an IC50 of 4.76 µg/ml for BALB/c and 81.92 µg/ml for C57BL/6.
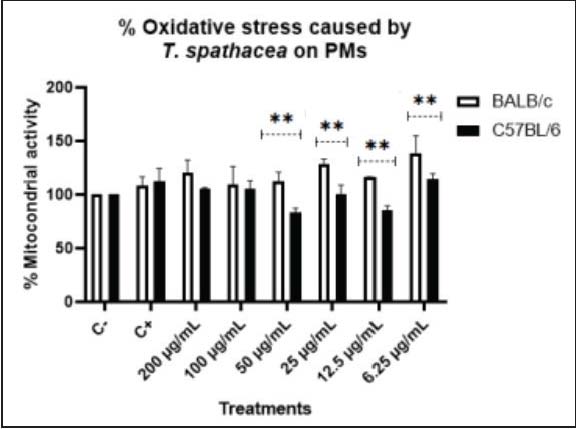
Fig. 1. Effects of the MET.s on mitochondrial activity in LPS-stimulated PMs. C+: positive control [LPS (100 ng/ml)], C-: negative control (DMEM 10% FBS).
A modest upward trend in mitochondrial activity was observed in BALB/c macrophages, although not statistically significant (Fig. 1). Flavonoids present in T. spathacea have been reported to attenuate inflammatory responses (Butnariu et al., 2022) by inhibiting cytokine secretion, which may suggest that MET.s exert mild immunomodulatory effects without compromising mitochondrial function.
In contrast, C57BL/6 macrophages showed lower mitochondrial activity, particularly at 50 and 12.5 µg/ml concentrations, compared with controls. This can be explained by their metabolic shift after stimulation, which can change to aerobic glycolysis to ensure energy production (Cai et al., 2023).
NO production
MET.s did not significantly alter NO production in PMs of either strain, except at the intermediate concentration of 50 µg/ml, where a significant difference between strains was observed, showing a p value of ≤0.0021. BALB/c macrophages displayed a more homogenous response, with no significant differences among treatments. In contrast, C57BL/6 macrophages showed decreased NO production, especially at 50 µg/ml.
It has been established that C57BL/6 macrophages generally produce higher levels of NO than BALB/c macrophages upon stimulation with LPS (Restrepo et al., 2025). Consistent with this, our LPS-treated positive controls showed higher NO production in C57BL/6 cells than in BALB/c cells (Fig. 2). Normally, this contrast is attributed to differences in arginine metabolism, where arginase activity predominates in BALB/c macrophages, whereas iNOS activation is favored in C57BL/6 macrophages (Soudi et al., 2013). Moreover, the stimulation exposure time also influences NO dynamics. For instance, higher NO levels are detected in BALB/c macrophages at 3 and 24 hours post-LPS stimulation, whereas C57BL/6 macrophages can only produce higher amounts of NO than BALB/c after 72 hours, consistent with their Th2- and Th1-biased immune profiles (Soudi et al., 2013).
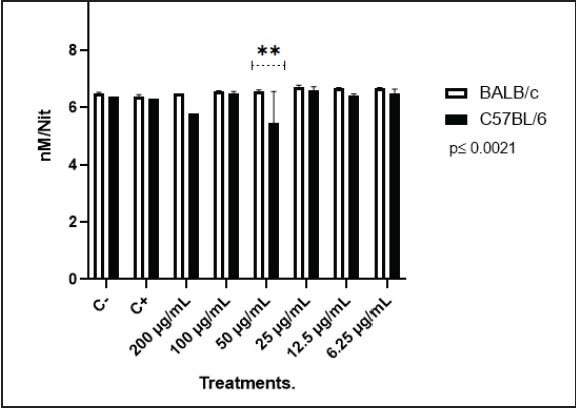
Fig. 2. NO production in PMs. C+: positive control [LPS (100 ng/ml)], C-: negative control (DMEM 10% FBS).
Notably, in this case, the NO production pattern was inverted compared with that is typically observed between strains, as MET.s appeared to exert a mild immunostimulatory effect on BALB/c macrophages while reducing NO production on C57BL/6 macrophages.
Cytokine gene expression in BALB/c PMs
Cytokine gene expression was assessed by qPCR under different treatments.
In LPS-stimulated macrophages, TGF-β expression was markedly elevated, even exceeding TNF-α (Fig. 3). Although LPS is a potent TLR4 activator that typically induces an M1 response with strong pro-inflammatory cytokine production (Martinez and Gordon, 2014), it can also trigger TGF-β/SMAD- mediated anti-inflammatory pathways, limiting tissue damage and promoting inflammation resolution (Zheng and Harris, 2021). This pattern may indicate an early balance between PM activation and regulation. On the other hand, under LPS + MET.s conditions, iNOS remained the most expressed gene, suggesting an additive or synergistic effect on pro-inflammatory signaling. This may result from enhanced TLR4 activation by the extract’s compounds or endotoxins, or from metabolic reprogramming towards aerobic glycolysis, which amplifies NOS2 expression (Corcoran and O’Neill, 2016).
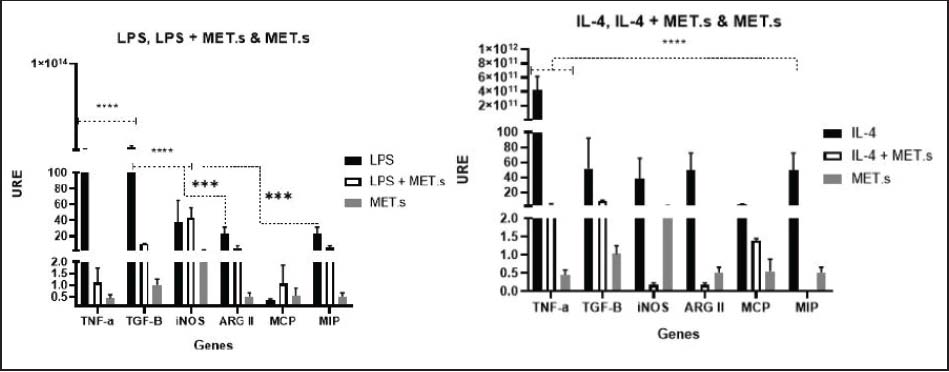
Fig. 3. Cytokine gene expression in BALB/c PMs under different treatments. URE: Relative Units of Expression.
Under IL-4 stimulation, a classic M2-polarizing cytokine, unexpectedly increased TNF-α expression compared to other genes, along with high variability between replicates (Fig. 3). These findings may reflect the presence of residual pro-inflammatory cues or basal pre-activation resulting from cell isolation, as previously described in primary cultures (Van den Bossche et al., 2015). IL-4 + MET.s co-stimulation predominantly induced TGF-β, followed by TNF-α. IL-4 signaling activates STAT6 and promotes M2 polarization, which is associated with tissue repair, matrix remodeling, and profibrotic mediator production, such as TGF-β (Zhu et al., 2023). The extract may potentiate these regulatory or fibrogenic pathways, possibly through nuclear receptor interactions or epigenetic modulation of IL-4 responsive promoters, generating a mixed response with both reparative and inflammatory components.
Exposure to MET.s alone induced iNOS expression, a hallmark of M1 activation responsible for NO synthesis from L-arginine, with potent microbicidal and pro-inflammatory effects (Zhang et al., 2021). This suggests that the extract contains metabolites capable of activating pro-inflammatory pathways, potentially via NF-κB or STAT1, as reported for certain plant-derived compounds that stimulate NO production in macrophages (Zhang et al., 2021).
Cytokine gene expression in C57BL/6 PMs
Under LPS+MET.s treatment, the expression of iNOS remained lower than with LPS alone, while ARGII and TGF-β were upregulated (Fig. 4). This tendency suggests that the extract could attenuate excessive pro-inflammatory signaling while promoting a metabolic shift towards M2. Although LPS is a potent TLR4 activator that typically induces an M1 response with strong pro-inflammatory cytokine production (Martinez and Gordon, 2014), it can also trigger TGF-β/SMAD-mediated anti-inflammatory pathways, limiting tissue damage and promoting inflammation resolution (Zheng and Harris, 2021), a pattern that may indicate an early balance between activation and regulation in PMs.
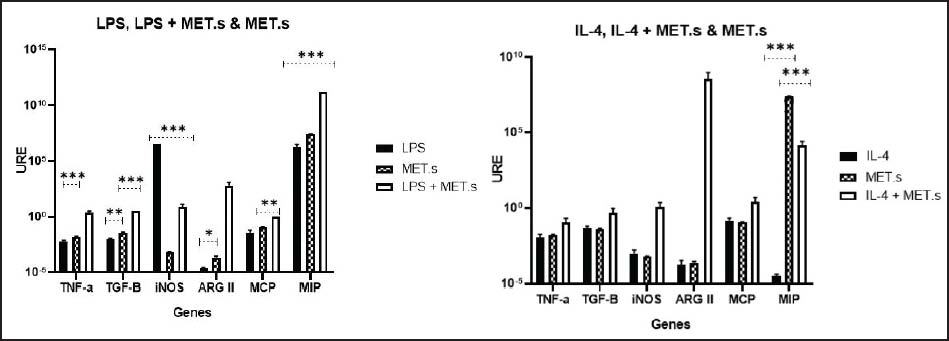
Fig. 4. Cytokine gene expression in C57BL/6 PMs under different treatments. URE: Relative Units of Expression.
Treatment with MET.s induced MIP overexpression in both LPS and IL-4, while other genes remained low. MCP reduction may indicate decreased monocyte-recruiting signaling (Singh et al., 2021), whereas MIP upregulation may indicate selective recruitment of regulatory or tissue-repair cells. MIP, particularly MIP-1α and MIP-2, orchestrate immune responses, leading to leukocyte migration through CCR1/CCR5 and CXCR2 signaling (Sindhu et al., 2020), mechanisms that could explain how MET.s favors a balanced chemotactic environment.
During IL-4 stimulation, a classic M2-polarizing cytokine, did not significantly alter the expression of the M2 marker TGF-β, but significantly increased ARGII in the IL-4+MET.s treatment, alongside a strong rise in MIP expression under both MET.s and IL-4+MET.s treatments (Fig. 4). It has been established that IL-4 signaling activates STAT6 and promotes M2 polarization, associated with tissue repair, matrix remodeling, and the production of profibrotic mediators such as TGF- β (Zhu et al., 2023). MET.s may potentiate these regulatory or fibrogenic pathways, possibly through nuclear receptor interactions or epigenetic modulation of IL-4-responsive promoters, generating a mixed response with both reparative and inflammatory components. This dual behavior indicates that MET.s exerts a context-dependent immunomodulatory effect, promoting homeostatic balance between activation and resolution mechanisms in macrophages.
These observations further support that the isolation/elicitation procedure may influence baseline activation programs and should be considered when interpreting polarization-related transcription patterns.
Study limitations
PMs were collected after a brief intraperitoneal exposure to LPS prior to lavage, which may prime TLR4-dependent programs and influence baseline gene expression. Therefore, our findings should be interpreted as MET.s-mediated modulation of LPS- or IL-4-conditioned responses in LPS-elicited PMs. Future studies should discern resident LPMs from recruited monocyte-derived macrophages and validate key findings in non-elicited resident cells. Additionally, endotoxin quantification and polymyxin B/TLR4 controls, along with further biochemical characterization of MET.s and protein/functional validation, are needed to exclude TLR4-driven artifacts and support translational claims.
Conclusion
The evidence suggests that MET.s acts as an immunomodulator of PMs, promoting a pro-nitro-oxidative response under pro-inflammatory conditions and a regulatory/pro-fibrogenic bias under alternative conditions without exhibiting cytotoxic effects. However, due to the chemical complexity of the extract, further analyses are required to elucidate its mechanism of action and potential therapeutic applications.
Acknowledgments
This paper is dedicated to all colleagues within the Immunology laboratory of the Faculty of Veterinary Medicine and Animal Science, to our family and friends.
Conflicts of interest
The authors have declared that no competing interests exist.
Funding
This study was supported by the 27th UANL Summer Program for Scientific and Technological Research (PROVERICYT-UANL). Code CT747-19 of the Universidad Autónoma de Nuevo León.
Authors’ contributions
D.L.CH.A: Conceptualization, Formal analysis, Resources, Writing - Original Draft, Writing - Review & Editing, Visualization. A.G.L.D: Conceptualization, Formal analysis, Resources, Writing - Original Draft, Writing - Review & Editing, Visualization K.F.V: Conceptualization, Formal analysis, Visualization. V.I.L.V: Conceptualization, Formal analysis, Visualization. G.H.V: Conceptualization, Formal analysis, Visualization R.A.P.H: Writing - Original Draft, Writing - Review & Editing A.S.D Conceptualization, Formal analysis, Visualization U.C.V: Conceptualization, Formal analysis, Resources, Writing - Original Draft, Writing - Review & Editing, Visualization, funding acquisition.
Data availability
The data supporting the findings of this study are publicly available and can be obtained without restriction from the institutional repository or through open-access platforms.
References
Arabpour, M., Saghazadeh, A. and Rezaei, N. 2021. Anti-inflammatory and M2 macrophage polarization-promoting effect of mesenchymal stem cell-derived exosomes. Intern. Immunopharm. 97, 107823.
Ardavín, C., Alvarez-Ladrón, N., Ferriz, M., Gutiérrez-González, A. and Vega-Pérez, A. 2023. Mouse tissue-resident peritoneal macrophages in homeostasis, repair, infection, and tumor metastasis. Ad. Sci. 10, 2206617.
Breda, J., Banerjee, A., Jayachandran, R., Pieters, J. and Zavolan, M. 2022. A novel approach to single-cell analysis reveals intrinsic differences in immune marker expression in unstimulated BALB/c and C57BL/6 macrophages. FEBS. Lett. 596, 2630–2643.
Butnariu, M., Quispe, C., Herrera-Bravo, J., Fernández-Ochoa, Á., Emamzadeh-Yazdi, S., Adetunji, C. O., Memudu, A. E., Otlewska, A., Bogdan, P., Antolak, H., Tamimi, K., Baghalpour, N., Bakhtiyari, J.M., Sen, S., Acharya, K., Segura-Carretero, A., De la Luz Cádiz-Gurrea, M., Lim, S.H.E., Pentea, M. and Sharifi-Rad, J. 2022. A review on tradescantia: phytochemical constituents, biological activities and health-promoting effects. Fro. In Bio. 27(6), 197.
Cai, S., Zhao, M., Zhou, B., Yoshii, A., Bugg, D., Villet, O., Sahu, A., Olson, G.S., Davis, J. and Tian, R. 2023. Mitochondrial dysfunction in macrophages promotes inflammation and suppresses repair after myocardial infarction. J. Cli. Inv. 133, 159498.
Chomczynski, P. 1993. Single-step method of total RNA isolation by guanidine–phenol extraction. En. Of Life Sci. 1–6.
Corcoran, S.E. and O’Neill, L.A. 2016. HIF1α and metabolic reprogramming in inflammation. J. Of C. Inv. 126(10), 3699–3707.
De Jesus, A., Pusec, C.M., Nguyen, T., Keyhani-Nejad, F., Gao, P., Weinberg, S.E. and Ardehali, H. 2022. Optimized protocol to isolate primary mouse peritoneal macrophage metabolites. STAR Prot. 3, 101668.
Han, Z., Shen, Y., Yan, Y., Bin, P., Zhang, M. and Gan, Z. 2025. Metabolic reprogramming shapes post-translational modification in macrophages. Mol. Asp. Med. 102, 101338.
Kushwaha, S., Kukshaal, M. and Patil, S.M. 2024. Review on Tradescantia spathacea (Medicinal Plant). J. For. Res. App. Sci. Biotech. 3, 104–115.
Lopes, L.E.S., Da Silva Barroso, S., Caldas, J.K.M., Vasconcelos, P.R., Canuto, K.M., Dariva, C., Santos, K.S., Severino, P., Cardoso, J.C., Souto, E.B. and Gomes, M.Z. 2024. Neuroprotective effects of Tradescantia spathacea tea bioactives in Parkinson’s disease: in vivo proof-of-concept. J. Trad. Comp. Med. 14, 435–445.
Lucía, C.P.A., Jacqueline, B.R., Alberto, B.R.L., David, B.A. and Beatriz, R.A. 2021. Actualized inventory of medicinal plants used in traditional medicine in Oaxaca, Mexico. J. Ethno. Ethnomed. 17(1), 1–15.
Mamani-Huanca, M., Muxel, S.M., Acuña, S.M., Floeter-Winter, L.M., Barbas, C. and López-Gonzálvez, A. 2021. Metabolomic reprogramming of C57BL/6-macrophages during early infection with L. amazonensis. Inter. J. Mol. Sci. 22, 6883.
Martinez, F.O. and Gordon, S. 2014. The evolution of our understanding of macrophages and translation of findings toward the clinic. Exp. Rev. Cli. Immuno. 11(1), 5–13.
Osmić, S., Begić, S. and Mićić, V. 2018. The effect of concentration of methanol as a solvent on the antioxidative activity of sage extract. Cham, Switzerland: Springer, , pp: 480–490.
Øynebråten, I., Tsatsanis, C., Ley, K., Org, K., Orecchioni, M., Ghosheh, Y. and Pramod, A.B. 2019. Present address: macrophage polarization: different gene signatures in M1(LPS+) vs. classically and M2(LPS-) vs. alternatively activated macrophages. Fron. Immuno. 1, 1084.
Restrepo, C.M., Llanes, A., Herrera, L., Ellis, E., Quintero, I. and Fernández, P.L. 2025. Baseline gene expression in BALB/c and C57BL/6 peritoneal macrophages influences but does not dictate their functional phenotypes. Exp. Bio. Med. 249(1), 10377.
Ross, E.A., Devitt, A. and Johnson, J.R. 2021. Macrophages: the good, the bad, and the gluttony. Fro. Immuno. 12, 708186.
Sindhu, S., Akhter, N., Wilson, A., Thomas, R., Arefanian, H., Al Madhoun, A., Al-Mulla, F. and Ahmad, R. 2020. MIP-1α expression induced by Co-stimulation of human monocytic cells with palmitate and TNF-α involves the TLR4-IRF3 pathway and is amplified by oxidative stress. Cells 9, 1799.
Singh, S., Anshita, D. and Ravichandiran, V. 2021. MCP-1: function, regulation, and involvement in disease. Inter. Immunopharm. 101, 107598.
Soudi, S., Zavaran-Hosseini, A., Muhammad Hassan, Z., Soleimani, M., Jamshidi Adegani, F. and Hashemi, S.M. 2013. Comparative study of the effect of LPS on the function of BALB/c and C57BL/6 peritoneal macrophages. Cell. J. 15, 45.
Sriwanthana, B., Treesangsri, W., Boriboontrakul, B., Niumsakul, S. and Chavalittumrong, P. 2007. In vitro effects of Thai medicinal plants on human lymphocyte activity. Songklanakarin. J. Sci. Tech. 29, 17–28.
Tan, J., Yap, W., Tan, S., Lim, Y. and Lee, S. 2014. Antioxidant content, antioxidant activity, and antibacterial activity of five plants from the Commelinaceae family. Antioxidants 3, 758–769.
Tan, J.B.L. and Kwan, Y.M. 2020. The biological activities of the spiderworts (Tradescantia). Food. Chem. 317, 126411.
Tu, P.C., Tseng, H.C., Liang, Y.C., Huang, G.J., Lu, T.L., Kuo, T.F. and Kuo, Y.H. 2019. Phytochemical investigation of Tradescantia albiflora and anti-inflammatory butenolide derivatives. Molecules 24, 3336.
Van Den Bossche, J., Baardman, J. and De Winther, M.P. 2015. Metabolic characterization of polarized M1 and M2 bone marrow-derived macrophages using real-time extracellular flux analysis. J. Of Vis. Exp. 105(1), 1579–1591.
Zhang, H., Zhang, Y., Liu, X., Elsabagh, M., Yu, Y., Peng, A., Dai, S. and Wang, H. 2021. L-Arginine inhibits hydrogen peroxide-induced oxidative damage and inflammatory response by regulating antioxidant capacity in ovine intestinal epithelial cells. It. J. Of A. Sci. 20(1), 1620–1632.
Zheng, G. and Harris, D.C.H. 2021. Editorial: tGF-β in human disease: friend or foe?. Fro. Cell Develop. Bio. 9, 739172.
Zhu, Y., Jin, X., Fu, N. and Li, J. 2023. Medrysone promotes corneal injury repair by promoting M2-like polarization of macrophages. BMC Oph. 23, 1–9.
Supplementary Materials
Table S1. Primers used for cytokine gene expression analysis.