




| Research Article | ||
Open Vet. J.. 2025; 15(6): 2682-2692 Open Veterinary Journal, (2025), Vol. 15(6): 2682-2692 Research Article Gastrointestinal parasite infection and the first molecular detection of strongyle infection in cattle of the Ayeyarwaddy Division, MyanmarBabi Kyi Soe1*, Apinya Arnuphapprasert2, Ana Huertas-López3, Toe Win Naing4, Khin Su Hlaing1 and Win Myint11Department of Livestock Breeding and Veterinary, Yangon, Myanmar 2Faculty of Veterinary Science, Rajamangala University of Technology Srivijaya, Songkhla, Thailand 3Investigación Multidisciplinar en Ciencias Veterinarias (IMCivet), Catholic University of Murcia (UCAM), Murcia, Spain 4Livestock Improvement Sector, De Heus Co. Ltd., Yangon, Myanmar *Correspondence to: Babi Kyi Soe. Department of Livestock Breeding and Veterinary, Yangon, Myanmar. Email: babikyisoe.vet [at] gmail.com Submitted: 28/02/2025 Revised: 10/05/2025 Accepted: 10/05/2025 Published: 30/06/2025 © 2025 Open Veterinary Journal
AbstractBackground: Gastrointestinal parasites (GI) are a group of pathogens that infect the digestive tract of a wide range of animals and cause significant infections in cattle worldwide. Nematodes at the order level Strongylida have the highest prevalence in livestock farming. In Myanmar, cattle production plays a critical role in the rural lifestyle. However, baseline data on GI parasitic infection in cattle from Myanmar remain scarce. Aim: This study is the first report to identify the most abundant GI parasites in cattle from this country using microscopic identification, followed by species identification of egg-positive samples through molecular methods targeting the internal transcribed spacer (ITS) region. Methods: A total of 219 cattle were involved in this cross-sectional study. Fecal flotation and formalin-ether centrifugal sedimentation techniques were used for conventional microscopic analysis. A fragment of the ITS gene was analyzed genetically in five samples identified as single-species infections. Results: Microscopy revealed an overall infection rate of 79.5% with intestinal parasites. Regarding results, strongyles were the most frequently detected parasites, followed by Eimeria spp. and Toxocara spp. Concretely, two species from the family Trichostrongylidae were found in the study area, Haemonchus contortus and Trichostrongylus colubriformis. Conclusion: This study provides the first molecular evidence of GI parasites in cattle in Myanmar, highlighting the high risk of parasitic infections in this area. Molecular analysis of five samples showed single-species infections: four with H. contortus and one with T. colubriformis, both likely widespread and dominant in Myanmar. These findings suggest that cattle may contribute to local transmission. The high prevalence underscores the necessity for sustained surveillance, implementation of effective control strategies, intersectoral collaboration between veterinary and public health authorities, and enhancement of public awareness to prevent and manage significant GI parasitic infections in Myanmar. Keywords: Cattle, Gastrointestinal parasites, Microscopy, Myanmar, Strongyle. IntroductionLivestock is vital to national economies and rural livelihoods, offering milk, meat, inputs for crop production, soil enrichment, and raw materials for industry and serving as a form of living insurance for farmers. The importance of dairy products and meat as essential protein sources for humans has led to an increase in cattle farming (Pighin et al., 2016). However, at the same time, significant adverse challenges, such as parasitic diseases, antimicrobial resistance, and climatic disasters, have also arisen (Soe et al., 2024). In Myanmar, over 70% of the population resides in rural areas, with more than 58% raising cattle, goats, and chickens as primary or supplementary sources of income for household expenses (Belton and Fang, 2022). The country has an estimated population of over 9.6 million draught cattle and approximately 129,000 crossbred dairy cattle. Cattle farming in Myanmar is threatened by numerous pathogenic threats, such as limited veterinary services, lack of knowledge, poor health practices, and the absence of preventive measures against parasitic infections. When a livestock population arises, pathogens and their diseases pose critical concerns for human and animal health. Either livestock or farm animals may act as sources of infections for humans or vice versa (Pradhan and Karanth, 2023), so the fight against these zoonoses must be carried out using a One Health approach (Sinclair, 2019). Regarding parasitic diseases, beyond causing high morbidity in both humans and animals, they result in considerable economic loss on a global scale associated with weight loss, reduced meat and milk production, abortions, and sterility (Lopes et al., 2016). In fact, herd mortality can exceed 40% due to either internal or external parasitic infections, particularly among young animals, and annual weight losses of 6–12 kg per animal have been reported (Sykes, 1994). While parasites are a highly diverse group of organisms, gastrointestinal (GI) parasites are of particular relevance in the context of cattle farming in Myanmar. GI parasites represent a major global burden, particularly affecting livestock health and productivity in tropical and subtropical regions, including Myanmar (Krecek and Waller, 2006; Pisarski, 2019). GI parasitism in ruminants occurs year-round, with higher intensity during the rainy season, as temperature and humidity have been mentioned as key factors that influence the development and survival of eggs, larvae, cysts, and oocysts in pastures (Terfa et al., 2023). Helminths, such as Strongyloides spp., which inhabit the small intestine or cecum, are the primary cause of GI parasitism in cattle, linking to sudden death in severe cases (Thamsborg et al., 2017). GI parasitism poses a significant threat to cattle farming in Myanmar, compounding existing challenges such as limited access to veterinary services, inadequate parasite control measures, and the absence of routine deworming practices. Understanding the epizootiology of parasitism is essential for sustainable parasitic control because it depends on interactions between the host, climate, management practices, and production conditions. However, the health status of the cattle population in the Ayeyarwaddy Division remains unclear, despite it being the second largest cattle-producing region in Myanmar, and to the author’s knowledge, no previous investigations have been conducted. The primary diagnostic method for parasitic infection is fecal microscopic examination. However, this technique requires the expertise of a skilled technician (Playford and Besier, 2024). Although microscopic examination of strongyle eggs in fecal samples is straightforward, the egg morphology does not permit species identification. For this purpose, internal transcribed spacer (ITS) regions in rDNA can be used to identify nematodes (Chilton, 2004). Genetic markers within the ITS enable accurate species-level identification of nematodes (Gasser et al., 1996; Newton et al., 1998). Recent studies of cattle GI parasites in other countries have revealed high prevalence rates and fluctuating diversity across them (Huang et al., 2014; Ahmed et al., 2015; Das et al., 2018; Income et al., 2021). Until now, updated baseline data on bovine GI parasites at the country level are still needed in Myanmar. The aims of the current study, therefore, are i) screen for major GI parasites in cattle fecal samples using a light microscopic technique and ii) the further molecular identification of nematode species based on ITS genetic markers. In addition, phylogenetic analyses between the isolates identified in the current study and other global isolates will be performed. These findings will serve as baseline data and contribute to developing effective strategies to tackle the health challenges associated with cattle farming practices in Myanmar. Materials and MethodsSampling area and farming characteristicsAcross-sectional study was conducted for approximately 4 months between August and November 2022. The sampling map, geographic coordinates, and livestock husbandry system of the Ayeyarwaddy Division are shown in Figure 1. The Ayeyarwaddy Division, located in the southern part of Myanmar, experiences tropical weather, with summer temperatures ranging from 24° to 36°C and winter temperatures from 19° to 31°C. Moreover, it is a low-lying delta region with extensive floodplains. The area is characterized by rivers, wetlands, and fertile agricultural land, making it one of Myanmar’s most important regions for rice production. Unlike the mountainous regions in the North and East of Myanmar, the Ayeyarwady Division has relatively flat topography. Livestock farming in the Ayeyarwaddy Division is predominantly practiced on a small scale and is integrated with crop production. The region is characterized by high humidity and seasonal flooding and is considered favorable for parasite transmission. Several challenges are commonly encountered, including limited access to veterinary care, irregular deworming practices, low input systems, and financial constraints, which contribute to suboptimal animal health and productivity. The study targeted 21 farms, of which each farm housed more than 10 cattle. A total of 219 cattle fecal samples were collected. These cattle were crossbreed (approximately two-third of population) and indigenous breeds, and they were raised using traditional method with semi-intensive farming practices when grazing pastures were available. Feeding practices included roughage, such as rice straw and natural grass and concentrated feeds such as rice bran and sesame cake chosen based on socioeconomic factors. Questionnaires surveyA questionnaire was designed to gather signalment data related to the history of the sampled animals. The questionnaire covered information on age (<1 year, 1–3 years, and >3 years), sex (male/ female), breed (crossbred/ indigenous), feeding system (grazing/ cut and carry), and anthelmintics usage (routine practice/ not regular). The age of the cattle was estimated using dentition, following the method described by Sten (1989). Cattle over 3 years old were classified as mature, as breeding (natural or artificial) is typically completed in Myanmar by this age. Accordingly, the cattle were categorized into three age groups, and all sampled cattle were identified using ear tags.
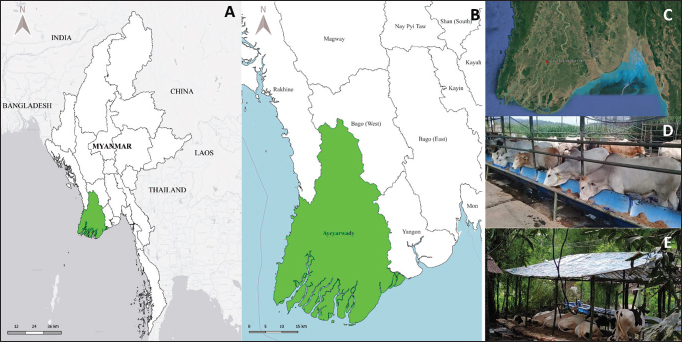
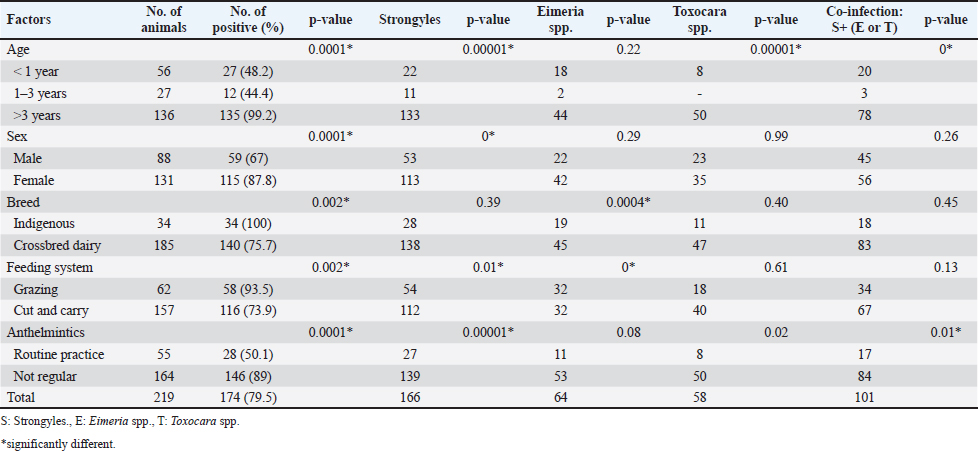
Fig. 1. (A, B, and C) Map and geographic coordinates of sampling sites in Ayeyarwaddy Division, southern part of Myanmar. (D and E). The figures demonstrate cattle livestock husbandry systems nearby residential areas and grazing pastures. The map was drawn by QGIS (version 3.36.3). The satellite image was modified from the Google Earth Website. Fecal sample collection, microscopic identification, and statistical analysisFecal samples were collected directly from the rectum using sterile plastic bags. Thereafter, the samples were placed in cool ice boxes and transported to the Laboratory of Molecular Detection, Livestock Upgrading Section, Mingaladon, Yangon, where fecal examination was conducted within 24 hours. For fecal flotation, 2 g of feces was mixed with 10 ml of saturated NaCl solution and transferred into a 15 ml corning tube by passing through a sterile cotton gauze. The tube was then filled with saturated NaCl until a convex meniscus formed at the top and was covered with a coverslip. After standing for 10–15 minutes, the coverslip was examined under a light microscope (Dryden et al., 2005). In formalin-ether centrifugal sedimentation, approximately 2 g of feces was mixed with 10 ml of 10% formalin and filtered through sterile cotton gauze into a 15 ml corning tube. The volume was adjusted to 10 ml by adding 10% formalin, followed by the addition of 2 ml of ether. The mixture was then shaken vigorously and centrifuged at 500 × g for 2 minutes. The sediment was then transferred to a glass slide, covered with a coverslip, and subsequently examined under a light microscope (Zajac and Conboy, 2012). Statistical analysisA Pearson chi-square test was conducted using SPSS version 28.0 (IBM Corp., NY, USA), to assess associations between the occurrence of infection, as determined by microscopic identification, and selected parameters: age, sex, breed, feeding system, and anthelmintic use. In addition, multiple regression analyses were conducted to evaluate the simultaneous effects of these variables on each parasite infection status. The analysis was performed using a 95% confidence interval, with a p-value <0.05 considered statistically significant. DNA extraction and polymerase chain reaction (PCR) amplificationDNA was extracted from five microscopically positive samples which showed a high number of fecal eggs using the QIAamp Fast DNAStool Mini Kit (Qiagen, Germany), according to the manufacturer’s protocol. Briefly, the ITS region of the nematode was amplified using the primers (ITSF 5′-CTCGCTCTCAGTGTTTTCGT-3') and (ITSR 5'-TTGACAAGTTAAGCAGCTTC-3'), as described by Gasser et al. (1996). PCR amplification was performed under the following conditions: initialization at 94°C for 5 minutes, 40 cycles of denaturation at 98°C for 10 seconds, annealing at 55°C for 15 seconds, extension at 68°C for 1 minute, and a final extension at 68°C for 5 minutes (Sato et al., 2014). The PCR products were analyzed using 1% agarose gel electrophoresis in 1× TBE buffer at 100 V for 30 minutes, and positive bands were checked using 1,000 bp DNA ladder (Thermo Fisher Scientific Inc., Waltham, MA). Sequencing and phylogenetic analysesThe amplified PCR products were sent to a commercial sequencing company (Macrogen, Seoul, South Korea). The resulting nucleotide sequences were then checked using a BLASTn search tool ( Haplotype networksHaplotype networks were constructed using 32 sequences of Trichostrongylus colubriformis, Oesophagostomum species, and Haemonchus species. The number of haplotypes was determined using DnaSP software (version 5.10.1). Representative haplotype sequences were manually arranged before visualization with the TCS network in PopART ( Ethical approvalNot needed for this study. ResultsMicroscopic identificationIn the present study, the overall rate of infection with intestinal parasites was 79.5% (174/219), indicating a high level of GI parasitic infection. The most common parasites were strongyles (95.4% (166/174), followed by Eimeria spp. 36.8% (64/174) and Toxocara spp. 33.3% (58/174). The coinfection of strongyles with either Eimeria spp. or Toxocara spp. was identified in 58% (101/174) of the cattle. The parasite stages detected via microscopy are shown in (Fig. 2), and the distribution of GI parasites relating to signalment data is shown in Table 1. According to the Pearson’s chi-square test, the prevalence of GI parasites was significantly higher in cattle older than 3 years (χ2 = 86.4; p = 0.0001), females (χ2 = 13.87; p = 0.0001), indigenous breeds (χ2 = 8.97; p = 0.02), animals with a feeding system based on grazing (χ2=9.36; p = 0.002), and those animals whose farmers did not routinely use anthelmintics (χ2 = 34.35; p = 0.0001). Molecular identification and phylogenetic analysesOnly five representative fecal samples were submitted for molecular amplification targeting the ITS gene, and amplicons were subsequently sent for Sanger sequencing. As a result, four sequences revealed 99.64% sequence identity with H. contortus sequence (accession no. AB908962) in ruminant fecal samples from Lao PDR. The remaining sequence showed 100% identity with T. colubriformis sequence (accession no. AB908960) from a ruminant fecal sample obtained from Lao PDR. The resulting five sequences were submitted to GenBank under accession number (PV069068-PV069072). Phylogenetic analyses were conducted to investigate closely related sequences (Fig. 3).
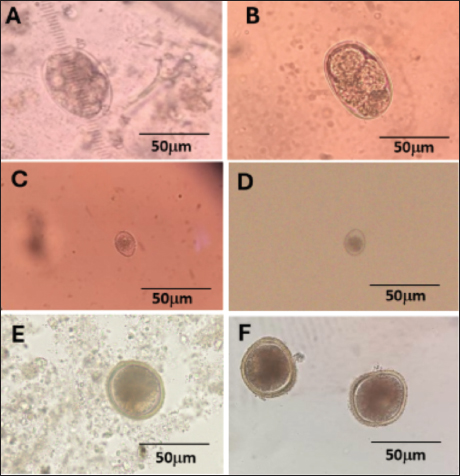
Fig. 2. GI parasite stages in cattle fecal sample in the present study. (A and B) Strongyles eggs, (C and D) Eimeria spp. oocyst, and (E and F) Toxocara spp. eggs observed under light microscope. A, C, E: sedimentation; B, D, F: flotation. Table 1. Distribution of GI parasites via microscopy.
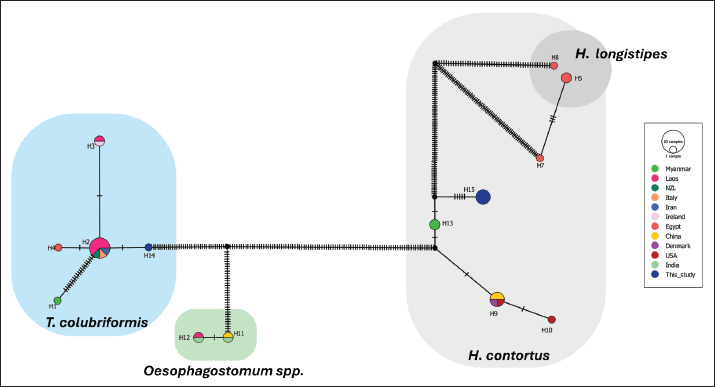
Haplotype networksHerein, a total of 15 identical haplotypes were identified in this study. The sequences from this study belonged to haplotype 14 (H14), representing T. colubriformis, and haplotype 15 (H15), representing H. contortus. Briefly, H14 was grouped with sequences from Myanmar, Laos, New Zealand, Italy, Iran, Ireland, and Egypt. Similarly, an identical H15 was clustered with Haemonchus spp. sequences from Egypt, Myanmar, China, Denmark, and the USA. Based on our analyses, no sequences from this study were related to Oesophagostomum species. Nucleotide variation was observed within each parasite group, particularly in Haemonchus spp. (Fig. 4).
Fig. 3. A phylogenetic analysis was performed using five sequences from this study and 28 reference sequences retrieved from GenBank, including two Oesophagostomum species, two Oesophagostomum columbianum, 11 T. colubriformis, and 13 H. contortus sequences. The maximum likelihood method, based on the TPM2+F+G4 model, was applied with 1,000 bootstrap replicates in IQTree software. Branch lengths in the tree represent the number of substitutions per site. Sequences from this study are highlighted in red. DiscussionUnderstanding the dynamics of parasitism is essential for efficient preventive measures in managing GI parasites (Cable et al., 2017). The relatively high prevalence rate (35.7%–86.9%) of GI parasitic infections in the livestock industry in Myanmar’s neighboring countries highlighted the need for public awareness and a treatment regime plan (Huang et al., 2014; Ahmed et al., 2015; Das et al., 2018; Income et al., 2021). To date, nematodes have been investigated as the most dominant GI parasites all over the world compared to trematodes and coccidia infection (Charlier et al., 2018). Although various GI parasites infect either livestock or farm animals, only strongyles, coccidia, flukes, and Moniezia spp. cause major GI problems, leading to chronic economic losses by reducing animal production rather than causing mortality (Jittapalapong et al., 2011). From GI nematodes, strongyles infection had been previously shown as the most common (Jittapalapong et al., 2011; Income et al., 2021). This study provides information about GI parasites in cattle farming areas in Ayeyarwaddy Division, Myanmar. The results revealed a high prevalence of GI parasites, with an overall rate of 79.5%. The different prevalence rates could be due to the presence or absence of some of the identified risk factors or the study design. For example, using more than one fecal sample from each individual, it is possible to improve diagnostic sensitivity, as single sampling for coprological techniques may result in an underestimation of the prevalence due to intermittent egg shedding of some parasites (Ferreira-Sá et al., 2024). In addition, cross-sectional study design does not assess the intensity and persistence of infections over time (Møller et al., 2003). In our study, the most abundant parasite stages were strongyle-type eggs 95.4% (166/174), followed by Eimeria spp. oocysts 36.8% (64/174) and Toxocara spp. eggs 33.3% (58/174). Co-infection of strongyles with either Eimeria spp. or Toxocara spp. was detected in 58% (101/174) of the cattle. The noticeable prevalence rate in the selected sampling division, which is situated near rivers, canals, and lakes, is consistent with previous findings showing that humidity and water sources play crucial roles in the parasite lifecycle (Jittapalapong et al., 2011). The infective stages of Toxocara spp. are shed in feces and require several days to weeks in the environment to embryonate and become infective. Therefore, high humidity and moist environments promote egg survival and embryonation, facilitating transmission through the ingestion of contaminated material (Mizgajska, 2001). In Eimeria, sporulation of oocysts occurs to become the infective stage, and sporulation is highly dependent on environmental conditions from which humid conditions favor the persistence and infectivity of oocysts (Sánchez et al., 2008). In strongyle nematodes, warm and humid environments with adequate moisture enhance larval development and migration onto vegetation, increasing the likelihood of host ingestion (Stromberg, 1997). In the sampling area, the Ayeyarwaddy River served as the main water reservoir for this division. Therefore, this river acts as a natural source for spreading these parasitic diseases, as it is located near villages where eggs from reservoir hosts can be shed (Vanhove et al., 2022). In this study, high GI parasitic infections, such as strongyles, Eimeria spp., and Toxocara spp. infection, might result from poor hygiene in housing pens, mixed crop-livestock systems, and contaminated environments, contributing to reinfection and high parasite burdens (Jittapalapong et al., 2011). Without proper sanitation, parasites are likely to pose a significant threat in the study area. Traditionally, cattle farming practices were commonly observed nearby residential areas (15–40 m) from the farm owner’s house; therefore, the distribution of infections appeared to be capable under poor hygienic behavior (Wangboon et al., 2024). Although strongyles are known to be primarily parasitized herbivores, sporadic human cases have been documented globally (Sato et al., 2010; Aung et al., 2017). Therefore, a broader systematic investigation of strongylosis is necessary given that free-roaming practices can increase the risk of parasitic diseases in humans. Intensive group housing further increases the risk of transmission within herds. Coccidiosis, caused by Eimeria spp., is a global intestinal disease in livestock that can cause anemia, diarrhea, and electrolyte imbalances (Jittapalapong et al., 2011). Eimeria species vary in pathogenicity, host specificity, and site of infection. Up to date, over 20 Eimeria spp. have been reported in cattle with most regarded as non-pathogenic or the animals have developed immunity after previous infection (Cornelissen et al., 1995). However, Eimeria bovis and Eimeria zuernii are known to be highly pathogenic in cattle (Munyua and Ngotho, 1990). Therefore, further molecular or morphological identification is necessary to distinguish between Eimeria species and evaluate their respective pathogenic potential in the study population. In the present study, Eimeria spp. were detected mainly in crossbred females over 3 years of age, indicating ongoing exposure and possible subclinical infection. In addition, the lack of regular deworming may have contributed to the persistence of environmental oocysts. Since adult cattle can act as reservoirs, there is a need for continued monitoring and parasite control across all age groups. Environmental contamination of potentially infectious sources, such as contaminated pastures and water reservoirs, can facilitate oocyst transmission to young animals. Infection with Eimeria is considered a risk factor when animals experience stress, such as transportation or underfeeding, which can weaken the immune system and result in the proliferation of Eimeria in the intestine (Chartier and Paraud, 2012). Toxocara spp. infections in cattle are uncommon; however, the ingestion of embryonated Toxocara eggs from contaminated environments leads to larval migration within the tissues of livestock animals (Ziegler and Macpherson, 2019). Among Toxocara spp., Toxocara vitulorum is a pathogenic GI nematode that infects cattle and buffalo in tropical and subtropical regions, where calves raised in contaminated environments are at increased risk of infection. Heavy infections in young calves may cause intestinal obstruction and, in severe cases, death, making this parasite clinically and economically important (Rast et al., 2013).
Fig. 4. Haplotype network (TCS network) of T. colubriformis, Haemonchus spp., and Osesophagostomum spp. sequences. The network illustrates the relationships among 15 identical haplotypes, with sequences from this study belonging to haplotype 14 (H14) for T. colubriformis and haplotype 15 (H15) for H. contortus (colored by dark blue). Regarding statistical analyses, GI infections appeared to be higher in older cattle, suggesting either prolonged exposure or different immune responses among the groups (Jittapalapong et al., 2011; Sirbu et al., 2020). Similar to our findings, a higher prevalence of GI parasitic infection in adult cattle has been reported by Income et al. (2021), while Raza et al. (2007) found younger cattle were more prone to intestinal helminth infections. The influence of sex on GI parasite infections in ruminants appears to be multifactorial and context-dependent. Although female ruminants may be more susceptible due to physiological stressors (Paul et al., 2020), the behaviors and management practices of male ruminants can also increase the risk of GI infections (Mpofu et al., 2020). However, no significant sex-related differences in GI parasitic infections have been reported (Income et al., 2021). Similar to Sayeed et al. (2024), indigenous cattle were significantly more infected with GI parasites than crossbred cattle in the present study. This could be due to the grazing habitat of all indigenous cattle in small-scale farming systems due to the limited pasture area in Myanmar, whereas crossbred dairy cattle are kept in a semi-intensive grazing system, which reduces prolonged exposure to parasitic infection in grazing pasture. Supporting this fact, sharing grazing pasture has been reported as a potential risk of parasitic infection in ruminants (Chuenpreecha et al., 2014). The lack of regular deworming practice was significantly associated with parasitic infection in the present study. GI parasitic infections were linked to farm management and deworming intervals. Most cattle in intensive farms followed a 6-month deworming schedule using either ivermectin, albendazole, or fenbendazole, a routine that was generally inadequate (Income et al., 2021). These anthelmintics are ineffective against protozoa parasites such as Eimeria spp. (Foreyt, 2013). Therefore, lesser deworming intervals with suitable drug usage, along with farmer training, routine surveillance, and prompt treatment after proper diagnosis, are recommended to reduce parasite transmission. In addition, the molecular identification of strongyles was conducted in five samples which showed single-species (four with H. contortus and one with T. colubriformis) infection. Previously, T. colubriformis and H. contortus had been reported to be widely distributed in different parts of Lao PDR, Thailand, Indonesia, and Pakistan (Sato et al., 2014; Purwati et al., 2017; Income et al., 2021; Ahmad et al., 2024). Therefore, our study revealed common findings of dominant strongyle species similar to the above-mentioned studies. These two species are supposed to be widely distributed and probably the most dominant GI parasites in Myanmar. Consequently, the capabilities of the cattle on local transmission of these parasitic infections should be contemplated. Haemonchus contortus spp. pose a significant challenge to ruminant production because of their high egg production, with females producing thousands of eggs daily (Biffa et al., 2006). Trichostrongylus colubriformis can cause villous atrophy, increased intestinal permeability, and protein loss, leading to diarrhea and reduced weight gain, which later result in poor feed conversion efficiency in infected animals (Greer et al., 2009). Therefore, we highlight the significant role of cattle as reservoirs of GI parasites in Myanmar, but also, it should be taken into account that other zoonotic parasites could be co-infecting these animals, even when we have not identified them (probably because of the limitations of the diagnostic techniques employed, such as the low sensitivity of the fecal microscopy and the low number of samples analyzed by molecular methods). To sum up, the high prevalence of GI parasitic infection in cattle in Myanmar highlights the importance of continuous surveillance and effective control measures and the need for collaboration between the veterinary and public health sectors. Since a cross-sectional study in specific areas may not represent the intensity and persistence of the infection overtime, a further longitudinal approach would be beneficial to evaluate the infection dynamics more accurately. ConclusionOverall, our study provides updated knowledge on the most abundant GI parasitic infection in cattle in the Ayeyarwaddy Division, Myanmar. In this study, the only GI parasites identified were strongyles, Eimeria spp., and Toxocara spp., H. contortus, and T. colubriformis were identified through molecular detection, and phylogenetic analysis revealed their genetic relationship to previously reported sequences in neighboring countries. Our findings highlight the importance of continuous surveillance and the development of effective control strategies for GI parasites in livestock. The high prevalence of parasitic infections in cattle underscores the need for further investigations across different regions of Myanmar, particularly to address the gaps in diagnostic techniques and enhance the understanding of parasite dynamics. Collaboration between the veterinary and public health sectors is essential for improving control measures and mitigating the impact of these infections. AcknowledgmentsWe appreciate the farm owners’ permission to collect samples from their animals. Conflict of interestThe authors declare that they have no conflicts of interest. FundingThis study was funded jointly by the authors. Authors contributionsB.K.S. conceptualized the idea of the study and designed the experiments. B.K.S., K.S.H., and T.W.N. collected the samples. B.K.S., K.S.H., and W.M. underwent laboratory investigations. B.K.S., A.A., and A.H.L. analyzed the data. B.K.S. wrote the original manuscript. B.K.S., A.H.L., and A.A. revised the manuscript. All authors have read and approved the final version of the manuscript. ReferencesAhmad, N., Khan, S.A., Majid, H.A., Ali, R., Ullah, R., Bari, A., Akbar, N.U. and Majid, A. 2024. Epidemiology and phylogeny of Haemonchus contortus through internal transcribed spacer 2 gene in small ruminants. Front. Vet. Sci. 11, 1380203. Ahmed, R., Biswas, P.K., Barua, M., Alim, M.A., Islam, K. and Islam, M.Z. 2015. Prevalence of gastrointestinal parasitism of cattle in Banskhali upazilla, Chittagong, Bangladesh. J. Adv. Vet. Anim. Res. 2, 484–488. Aung, W.P.P., Htoon, T.T., Tin, H.H., Sanpool, O., Jongthawin, J., Sadaow, L., Phosuk, I., Ropai, R., Intapan, P.M. and Maleewong, W. 2017. First molecular identifications of Necator americanus and Ancylostoma ceylanicum infecting rural communities in lower Myanmar. Am. J. Trop. Med. Hyg. 96, 214. Belton, B. and Fang, P. 2022. Livestock, capture fisheries, and aquaculture in Myanmar: status and recent trends. Washington, DC: International Food Policy Research Institute, vol. 20. Biffa, D., Jobre, Y. and Chakka, H. 2006. Ovine helminthosis, a major health constraint to productivity of sheep in Ethiopia. Anim. Heal. Res. Rev. 7, 107–118. Cable, J., Barber, I., Boag, B., Ellison, A.R., Morgan, E.R., Murray, K., Pascoe, E.L., Sait, S.M., Wilson, A.J. and Booth, M. 2017. Global change, parasite transmission and disease control: lessons from ecology. Philo. Trans. R. Soc. B. 372, 20160088. Charlier, J., Thamsborg, S.M., Bartley, D.J., Skuce, P.J., Kenyon, F., Geurden, T., Hoste, H., Williams, A.R., Sotiraki, S., Höglund, J. and Chartier, C. 2018. Mind the gaps in research on the control of gastrointestinal nematodes of farmed ruminants and pigs. Transbound. Emerg. Dis. 65, 217–234. Chartier, C. and Paraud, C. 2012. Coccidiosis due to Eimeria in sheep and goats, a review. Small Rumin. Res. 103, 84–92. Chilton, N.B. 2004. The use of nuclear ribosomal DNA markers for the identification of bursate nematodes (order Strongylida) and for the diagnosis of infections. Anim. Health Res. Rev. 5, 173–187. Chuenpreecha, S., Semaming, Y., Pilachai, R., Kummee, P. and Sittijunda, S. 2014. Prevalence of gastro-intestinal strongyles in native beef cattle under smallholder management condition in Udon Thani, Thailand. Khon Kaen Agr. J. 42, 11–15. Cornelissen, A.W., Verstegen, R., van den Brand, H., Perie, N.M., Eysker, M., Lam, T.J. and Pijpers, A. 1995. Anobservational study of Eimeria species in housed cattle on Dutch dairy farms. Vet. Parasitol. 56, 7–16. Das, M., Deka, D.K., Sarmah, A.K., Sarmah, P.C. and Islam, S. 2018. Gastrointestinal parasitic infections in cattle and swamp buffalo of Guwahati, Assam, India. Indian J. Anim. Res. 52, 1732–1738. Dryden, M.W., Payne, P.A., Ridley, R. and Smith, V. 2005. Comparison of common fecal flotation techniques for the recovery of parasite eggs and oocysts. Vet. Ther. 6, 15–28. Ferreira-Sá, L.C., Machado, E.R., Gurgel-Gonçalves, R. and Abad-Franch, F. 2024. Disentangling the effects of intermittent faecal shedding and imperfect test sensitivity on the microscopy-based detection of gut parasites in stool samples. PLoS Negl. Trop. Dis. 18, e0012719. Foreyt, W.J. 2013. Veterinary parasitology reference manual. Hoboken, NY: John Wiley & Sons. Gasser, R.B., Stewart, L.E. and Speare, R. 1996. Genetic markers in ribosomal DNA for hookworm identification. Acta Trop. 62, 15–21. Greer, A.W., Sedcole, R.J., Jay, N.P., McAnulty, R.W., Green, R.S., Stankiewicz, M. and Sykes, A.R. 2009. Protein supply influences the nutritional penalty associated with the development of immunity in lambs infected with Trichostrongylus colubriformis. Animal 3, 437–445. Huang, C.C., Wang, L.C., Pan, C.H., Yang, C.H. and Lai, C.H. 2014. Investigation of gastrointestinal parasites of dairy cattle around Taiwan. J. Microbiol. Immunol. Infect. 47, 70–74. Income, N., Tongshoob, J., Taksinoros, S., Adisakwattana, P., Rotejanaprasert, C., Maneekan, P. and Kosoltanapiwat, N. 2021. Helminth infections in cattle and goats in Kanchanaburi, Thailand, with focus on strongyle nematode infections. Vet. Sci. 8, 324. Jittapalapong, S., Sangwaranond, A., Nimsuphan, B., Inpankaew, T., Phasuk, C., Pinyopanuwat, N., Chimnoi, W., Kengradomkij, C., Arunwipat, P. and Anakewith, T. 2011. Prevalence of gastro-intestinal parasites of dairy cows in Thailand. J. Agric. Nat. Res. 45, 40–45. Krecek, R.C. and Waller, P.J. 2006. Towards the implementation of the “basket of options” approach to helminth parasite control of livestock: emphasis on the tropics/subtropics. Vet. Parasitol. 139, 270–282. Lopes, L.B., Nicolino, R., Capanema, R.O., Oliveira, C.S.F., Haddad, J.P.A. and Eckstein, C. 2016. Economic impacts of parasitic diseases in cattle. CABI Rev. 10(51), 1–10. Mizgajska, H. 2001. Eggs of Toxocara spp. in the environment and their public health implications. J. Helminthol. 75, 147–151. Munyua, W.K. and Ngotho, J.W. 1990. Prevalence of Eimeria species in cattle in Kenya. Vet. Parasitol. 35, 163–168. Mpofu, T.J., Nephawe, K.A. and Mtileni, B. 2020. Prevalence of gastrointestinal parasites in communal goats from different agro-ecological zones of South Africa. Vet. World. 13, 26. Møller, A.P., Erritzøe, J. and Saino, N. 2003. Seasonal changes in immune response and parasite impact on hosts. Am. Nat. 161, 657–671. Newton, L.A., Chilton, N.B., Beveridge, I., Hoste, H., Nansen, P. and Gasser, R.B. 1998. Genetic markers for strongylid nematodes of livestock defined by PCR-based restriction analysis of spacer rDNA. Acta Trop. 69, 1–15. Paul, B.T., Jesse, F.F.A., Chung, E.L.T., Che’Amat, A. and Mohd Lila, M.A. 2020. Risk factors and severity of gastrointestinal parasites in selected small ruminants from Malaysia. Vet. Sci. 7, 208. Pighin, D., Pazos, A., Chamorro, V., Paschetta, F., Cunzolo, S., Godoy, F., Messina, V., Pordomingo, A. and Grigioni, G. 2016. A contribution of beef to human health: a review of the role of the animal production systems. Sci. World J. 1, 1–10. Pisarski, K. 2019. The global burden of disease of zoonotic parasitic diseases: top 5 contenders for priority consideration. Trop. Med. Infect. Dis. 4, 44. Playford, M.C. and Besier, R.B. 2024. Gastrointestinal nematode parasites of grazing ruminants: a comprehensive literature review of diagnostic methods for quantifying parasitism, larval differentiation and measuring anthelmintic resistance. N. Z. Vet. J. 73(3), 149–164. Pradhan, A.K. and Karanth, S. 2023. Zoonoses from animal meat and milk. In Present knowledge in food safety. Cambridge, MA: Academic Press, pp: 394–411. Purwati, E., Putra, M.S., Priyowidodo, D., Ribeiro da Silva, L.M. and Humaidah, H. 2017. Site distribution and identification of parasitic strongyle from cattle in Central Java, Indonesia. Asian Pac. J. Trop. Dis. 7, 539–543. Rast, L., Lee, S., Nampanya, S., Toribio, J.A.L., Khounsy, S. and Windsor, P.A. 2013. Prevalence and clinical impact of Toxocara vitulorum in cattle and buffalo calves in northern Lao PDR. Trop. Anim. Health Prod. 45, 539–546. Raza, M.A., Iqbal, Z., Jabbar, A. and Yaseen, M. 2007. Point prevalence of gastrointestinal helminthiasis in ruminants in southern Punjab, Pakistan. J. Helminthol. 81, 323–328. Sánchez, R.O., Romero, J.R. and Founroge, R.D. 2008. Dynamics of Eimeria oocyst excretion in dairy calves in the Province of Buenos Aires (Argentina), during their first 2 months of age. Vet. Parasitol. 151, 133–138. Sato, M., Sanguankiat, S., Yoonuan, T., Pongvongsa, T., Keomoungkhoun, M., Phimmayoi, I., Boupa, B., Moji, K. and Waikagul, J. 2010. Copro-molecular identification of infections with hookworm eggs in rural Lao PDR. Trans. R. Soc. Trop. Med. Hyg. 104, 617–622. Sato, M.O., Sato, M., Chaisiri, K., Maipanich, W.,Yoonuan, T., Sanguankiat, S., Pongvongsa, T., Boupha, B., Moji, K. and Waikagul, J. 2014. Nematode infection among ruminants in monsoon climate (Ban-Lahanam, Lao PDR) and its role as food-borne zoonosis. Rev. Bras. Parasitol. Vet. 23, 80–84. Sayeed, M.A., Ungar, L., Chowdhury, Y.H., Bari, M.S., Rahman, M.M., Anwer, M.S. and Hoque, M.A. 2024. Gastrointestinal parasitosis in cattle: unveiling the landscape across diverse production systems in Bangladesh. Vet. Med. Sci. 10, e1325. Sirbu, C.B., Imre, K., Darabus, G., Suici, T., Mates, B. and Morariu, S. 2020. Prevalence of gastrointestinal parasitic infections in cattle and sheep in two regions of Romania. Turkish J. Vet. Anim. Sci. 44, 581–587. Sten, S.A. 1989. A method of age determination on archaeological and modern cattle (Bos taurus) by counting of tooth annuli in the cementum. Laborativ. Arkeol. 5, 195–199. Stromberg, B.E. 1997. Environmental factors influencing transmission. Vet. Parasitol. 72, 247–264. Soe, B.K., Naing, T.W., Mon, S.L.Y., Nway, N.C. and Sato, H. 2024. First molecular evidence of Anaplasma marginale infection in naturally infected cattle in Myanmar with severe hemolytic anemia. Adv. Anim. Vet. Sci. 12, 559–565. Sykes, A.R. 1994. Parasitism and production in farm animals. Anim. Sci. 59, 155–172. Terfa, W., Kumsa, B., Ayana, D., Maurizio, A., Tessarin, C. and Cassini, R. 2023. Epidemiology of gastrointestinal parasites of cattle in three districts in Central Ethiopia. Animals 13, 285. Thamsborg, S.M., Ketzis, J., Horii, Y. and Matthews, J.B. 2017. Strongyloides spp. infections of veterinary importance. Parasitology 144, 274–284. Trifinopoulos, L.T., Nguyen, A., von Haeseler, B.Q. and Minh, W. 2016. IQ-Tree A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 44, 232–235. Vanhove, M.P., Kmentová, N., Luus-Powell, W.J., Netherlands, E.C., de Buron, I. and Barger, M.A. 2022. A snapshot of parasites in tropical and subtropical freshwater wetlands: modest attention for major players. In Fundamentals of tropical freshwater wetlands. Amsterdam, The Netherlands: Elsevier, pp: 417–485. Wangboon, C., Martviset, P., Jamklang, M., Chumkiew, S., Penkhrue, W., Rangdist, S., Jirojwong, R., Phadungsil, W., Chantree, P., Grams, R. and Krenc, D. 2024. Microscopic and molecular epidemiology of gastrointestinal nematodes in dairy and beef cattle in Pak Chong district, Nakhon Ratchasima province, Thailand. Vet. World. 17(5), 1035–1043. Zajac, A.M. and Conboy, G.A. 2012. Fecal examination for the diagnosis of parasitism. Vet. Clin. Parasitol. 8, 72–73. Ziegler, M.A. and Macpherson, C.N. 2019. Toxocara and its species. CABI Reviews, pp: 1–27. | ||

| How to Cite this Article |
| Pubmed Style Soe BK, Arnuphapprasert A, Huertas-lópez A, Naing TW, Hlaing KS, Myint W. Gastrointestinal parasite infection and the first molecular detection of strongyle infection in cattle of the Ayeyarwaddy Division, Myanmar. Open Vet. J.. 2025; 15(6): 2682-2692. doi:10.5455/OVJ.2025.v15.i6.37 Web Style Soe BK, Arnuphapprasert A, Huertas-lópez A, Naing TW, Hlaing KS, Myint W. Gastrointestinal parasite infection and the first molecular detection of strongyle infection in cattle of the Ayeyarwaddy Division, Myanmar. https://www.openveterinaryjournal.com/?mno=245127 [Access: January 25, 2026]. doi:10.5455/OVJ.2025.v15.i6.37 AMA (American Medical Association) Style Soe BK, Arnuphapprasert A, Huertas-lópez A, Naing TW, Hlaing KS, Myint W. Gastrointestinal parasite infection and the first molecular detection of strongyle infection in cattle of the Ayeyarwaddy Division, Myanmar. Open Vet. J.. 2025; 15(6): 2682-2692. doi:10.5455/OVJ.2025.v15.i6.37 Vancouver/ICMJE Style Soe BK, Arnuphapprasert A, Huertas-lópez A, Naing TW, Hlaing KS, Myint W. Gastrointestinal parasite infection and the first molecular detection of strongyle infection in cattle of the Ayeyarwaddy Division, Myanmar. Open Vet. J.. (2025), [cited January 25, 2026]; 15(6): 2682-2692. doi:10.5455/OVJ.2025.v15.i6.37 Harvard Style Soe, B. K., Arnuphapprasert, . A., Huertas-lópez, . A., Naing, . T. W., Hlaing, . K. S. & Myint, . W. (2025) Gastrointestinal parasite infection and the first molecular detection of strongyle infection in cattle of the Ayeyarwaddy Division, Myanmar. Open Vet. J., 15 (6), 2682-2692. doi:10.5455/OVJ.2025.v15.i6.37 Turabian Style Soe, Babi Kyi, Apinya Arnuphapprasert, Ana Huertas-lópez, Toe Win Naing, Khin Su Hlaing, and Win Myint. 2025. Gastrointestinal parasite infection and the first molecular detection of strongyle infection in cattle of the Ayeyarwaddy Division, Myanmar. Open Veterinary Journal, 15 (6), 2682-2692. doi:10.5455/OVJ.2025.v15.i6.37 Chicago Style Soe, Babi Kyi, Apinya Arnuphapprasert, Ana Huertas-lópez, Toe Win Naing, Khin Su Hlaing, and Win Myint. "Gastrointestinal parasite infection and the first molecular detection of strongyle infection in cattle of the Ayeyarwaddy Division, Myanmar." Open Veterinary Journal 15 (2025), 2682-2692. doi:10.5455/OVJ.2025.v15.i6.37 MLA (The Modern Language Association) Style Soe, Babi Kyi, Apinya Arnuphapprasert, Ana Huertas-lópez, Toe Win Naing, Khin Su Hlaing, and Win Myint. "Gastrointestinal parasite infection and the first molecular detection of strongyle infection in cattle of the Ayeyarwaddy Division, Myanmar." Open Veterinary Journal 15.6 (2025), 2682-2692. Print. doi:10.5455/OVJ.2025.v15.i6.37 APA (American Psychological Association) Style Soe, B. K., Arnuphapprasert, . A., Huertas-lópez, . A., Naing, . T. W., Hlaing, . K. S. & Myint, . W. (2025) Gastrointestinal parasite infection and the first molecular detection of strongyle infection in cattle of the Ayeyarwaddy Division, Myanmar. Open Veterinary Journal, 15 (6), 2682-2692. doi:10.5455/OVJ.2025.v15.i6.37 |

